Первичное продуцирование представляет собой синтез органического вещества в экосистемах. Весь процесс фотосинтеза в упрощенном виде можно записать следующим образом (Алимов, 1989):
![]() .
.
При расщеплении воды образуется газообразный кислород, из двуокиси углерода и восстановленного из воды водорода синтезируются углеводы и вновь образуется вода. В темноте фотосинтез прекращается, прекращается и потребление ![]() из среды и выделение эквивалентного количества кислорода. Процессы дыхания у растений в темноте идут с той же скоростью, что и на свету. Поэтому, сравнивая результаты жизнедеятельности водного сообщества на свету и в темноте, можно рассчитать значение первичной продукции. Как видно из уравнения, для синтеза органического вещества из двуокиси углерода и воды необходим источник энергии - свет. Утилизация энергии света является характерной особенностью фотосинтеза.
из среды и выделение эквивалентного количества кислорода. Процессы дыхания у растений в темноте идут с той же скоростью, что и на свету. Поэтому, сравнивая результаты жизнедеятельности водного сообщества на свету и в темноте, можно рассчитать значение первичной продукции. Как видно из уравнения, для синтеза органического вещества из двуокиси углерода и воды необходим источник энергии - свет. Утилизация энергии света является характерной особенностью фотосинтеза.
Первичная продукция в мелководных водоемах создается преимущественно фитопланктоном, перифитоном и макрофитами. В океанах и морях основной вклад в образование первичной продукции вносит фитопланктон. Фитопланктонное сообщество испытывает чрезвычайно разнообразные изменения условий среды, которые влияют самым непосредственным образом на скорость фотосинтеза и рост микроводорослей. Аккуратное рассмотрение и описание процессов первичного продуцирования должно учитывать многие физические и физико-химические факторы внешней среды, структурные особенности фитопланктонного сообщества, процессы, протекающие на уровне отдельной клетки.
Модели, в основу которых положена жизнедеятельность планктонных популяций, при внесении незначительных изменений можно использовать и для моделирования других сообществ в водных экосистемах, таких как макрофиты, перифитон и бентосные водоросли. Динамика биомассы фитопланктона в единице объема воды определяется, как правило, процессами роста, естественной смертности, выедания организмами высших трофических уровней, притока со стоком рек, с течениями и оттока в другие водоемы (или другие части водной экосистемы). Направление (рост или уменьшение) и скорость изменения биомассы зависит от соотношения скоростей различных процессов.
Характерные особенности процесса фотосинтеза состоят в том, что его световые и температурные оптимумы у разных видов фитопланктона различны. Для описания влияния температуры на скорость первичного продуцирования используют разнообразные эмпирические зависимости, например такие (Йоргенсен, 1985):
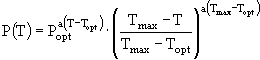 ; (1)
; (1)
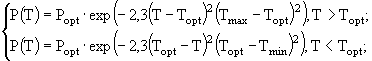 (2)
(2)
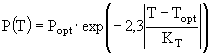 ; (3)
; (3)
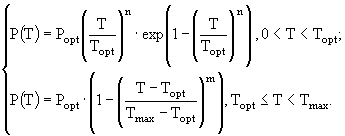 (4)
(4)
Здесь ![]() - интенсивность фотосинтеза при температуре T;
- интенсивность фотосинтеза при температуре T; ![]() - интенсивность фотосинтеза при оптимальной температуре
- интенсивность фотосинтеза при оптимальной температуре ![]() ;
; ![]() и
и ![]() - соответственно максимальная и минимальная температуры, при которых происходит фотосинтез; a,
- соответственно максимальная и минимальная температуры, при которых происходит фотосинтез; a, ![]() , m, n - эмпирические константы. Уравнение (3) несколько проще всех остальных, но оно обладает одним существенным недостатком - оно получено в предположении о том, что кривая зависимости скорости роста водорослей от температуры симметрична относительно точки оптимума, что не вполне согласуется с данными наблюдений. В ряде случаев применяют еще более простые соотношения, например, такое:
, m, n - эмпирические константы. Уравнение (3) несколько проще всех остальных, но оно обладает одним существенным недостатком - оно получено в предположении о том, что кривая зависимости скорости роста водорослей от температуры симметрична относительно точки оптимума, что не вполне согласуется с данными наблюдений. В ряде случаев применяют еще более простые соотношения, например, такое:
![]() , (5)
, (5)
где ![]() - интенсивность фотосинтеза при 20°С;
- интенсивность фотосинтеза при 20°С; ![]() - эмпирическая константа, немного большая единицы (обычно считают, что
- эмпирическая константа, немного большая единицы (обычно считают, что ![]() ). Это уравнение хорошо согласуется с экспериментальными данными только в определенных температурных интервалах. Однако его важным преимуществом является то, что оно содержит всего два эмпирических параметра.
). Это уравнение хорошо согласуется с экспериментальными данными только в определенных температурных интервалах. Однако его важным преимуществом является то, что оно содержит всего два эмпирических параметра.
Зависимость между скоростью первичного продуцирования и освещенностью может быть описана уравнением Михаэлиса-Ментен:
![]() , (6)
, (6)
где ![]() - скорость роста (или скорость первичного продуцирования);
- скорость роста (или скорость первичного продуцирования); ![]() - максимальная скорость роста;
- максимальная скорость роста; ![]() - освещенность;
- освещенность; ![]() - константа полунасыщения по освещенности, то есть такая величина I, при которой
- константа полунасыщения по освещенности, то есть такая величина I, при которой ![]() . При освещенности выше пороговой
. При освещенности выше пороговой ![]() , начиная с которой происходит угнетение фотосинтеза, вместо выражения (6) можно записать, например, такое (Йоргенсен, 1985):
, начиная с которой происходит угнетение фотосинтеза, вместо выражения (6) можно записать, например, такое (Йоргенсен, 1985):
![]() . (7)
. (7)
Значения всех констант (![]() ,
, ![]() ,
, ![]() ) зависят от адаптации к освещенности и температуры. Обычно для этого используют простейшие (как правило линейные) функциональные зависимости, содержащие небольшое число параметров.
) зависят от адаптации к освещенности и температуры. Обычно для этого используют простейшие (как правило линейные) функциональные зависимости, содержащие небольшое число параметров.
Помимо световых и температурных адаптаций темп роста ![]() зависит также и от концентраций биогенных веществ. Химический состав клеток фитопланктона непостоянен и до некоторой степени отражает содержание тех или иных веществ в воде. Рост может лимитироваться более чем одним биогенным веществом. Фактически все модели, использующие описание скоростей роста популяции или сообщества растительных организмов, основываются либо на принципе Либиха, либо на законе Митчерлиха (Страшкраба, Гнаук, 1989). В первом случае общая скорость моделируемого процесса определяется минимальной из скоростей потребления биогенных элементов, во втором - некоторой функциональной комбинацией скоростей потребления отдельных питательных элементов. Простейшим и, вместе с тем, наиболее часто применяемым в практике математического моделирования способом использования закона Либиха в системах дифференциальных уравнений для относительной скорости роста
зависит также и от концентраций биогенных веществ. Химический состав клеток фитопланктона непостоянен и до некоторой степени отражает содержание тех или иных веществ в воде. Рост может лимитироваться более чем одним биогенным веществом. Фактически все модели, использующие описание скоростей роста популяции или сообщества растительных организмов, основываются либо на принципе Либиха, либо на законе Митчерлиха (Страшкраба, Гнаук, 1989). В первом случае общая скорость моделируемого процесса определяется минимальной из скоростей потребления биогенных элементов, во втором - некоторой функциональной комбинацией скоростей потребления отдельных питательных элементов. Простейшим и, вместе с тем, наиболее часто применяемым в практике математического моделирования способом использования закона Либиха в системах дифференциальных уравнений для относительной скорости роста ![]() является следующий:
является следующий:
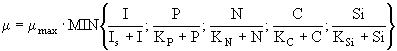 , (8)
, (8)
где ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() - соответственно константы полунасыщения по освещенности, фосфору и азоту, углероду и кремнию; P, N, C, Si - концентрации фосфора, азота, углерода и кремния; I - интенсивность освещенности. При использовании же закона Митчерлиха для описания зависимости относительной скорости роста
- соответственно константы полунасыщения по освещенности, фосфору и азоту, углероду и кремнию; P, N, C, Si - концентрации фосфора, азота, углерода и кремния; I - интенсивность освещенности. При использовании же закона Митчерлиха для описания зависимости относительной скорости роста ![]() фитопланктона от совокупности лимитирующих биогенных элементов и освещенности, как правило, используют мультипликативную зависимость:
фитопланктона от совокупности лимитирующих биогенных элементов и освещенности, как правило, используют мультипликативную зависимость:
![]() . (9)
. (9)
Следует отметить, что исходные формулировки принципов лимитирования как Либих, так и Митчерлих относят к популяциям одного вида. В то же время на практике эти принципы часто используются в применении к многовидовым сообществам. Концепция лимитирующих факторов в применении к сообществам фитопланктонных организмов теоретически и экспериментально исследована в работе (Левич и др., 1997; Левич и др., 1997).
Первые модели первичной продукции и роста микроводорослей имели очень простую структуру и создавались для оценки первичной продукции по известному распределению хлорофилла и интенсивности освещенности (Ryther, 1956). Фотосинтетические характеристики фитопланктона меняются с глубиной, зависят от времени суток, сезона, типа водоема и его географического положения. Всего этого первые модели никак не учитывали, из-за своей простоты. Постепенно шло усложнение моделей. Так, например, зависимость интенсивности фотосинтеза от глубины рассматривалась в моделях (Laws, Bannister, 1980; Kiefer, Mitchell, 1983; Smith et al., 1989). В дальнейшем начали учитывать влияние на рост фитопланктона температуры, концентрации биогенных элементов, отдельных биохимических процессов (Kiefer, Cullen, 1991; Sakshaug et al., 1991). Другие модели предусматривали учет влияния спектрального состава освещенности (Семовский, 1999; Семовский, 1999; Sathyendranath et al., 1989; Morel, 1991; Platt, Sathyendranath, 1991) и продолжительности светового дня (Sakshaug et al., 1991). В некоторых моделях также учитывалась внутрисуточная изменчивость освещенности (Wroblewski, 1977; Platt et al., 1990; Platt, Sathyendranath, 1991). Большинство из этих моделей рассматривают влияние на фотосинтез и первичную продукцию не более двух факторов среды. Например, в работах (Kiefer, Mitchell, 1983) и (Wroblewski, Richman, 1987) для моделирования роста фитопланктона в качестве внешних переменных рассматриваются фотосинтетически активная радиация (ФАР) и концентрации биогенных элементов, а в работе (Sakshaug et al., 1991) учитываются спектральный состав света и продолжительность светового дня (фотопериод). Модель, предложенная в работе (Smith et al., 1989), является более сложной и в ней анализируется совместное воздействие трех внешних факторов: спектральных характеристик освещенности, продолжительности светлого периода суток, а также внутрисуточных изменений интенсивности солнечной радиации. Еще более развитой является модель (Kiefer, Cullen, 1991). В качестве внешних переменных в модели используется ФАР, фотопериод, биогенные элементы и температура воды, но, в то же время, не учитываются спектральные особенности падающей радиации и ее внутрисуточная изменчивость.
Основой для изучения и количественного описания влияния интенсивности света на процесс фотосинтеза являются экспериментальные данные, полученные в лабораторных и естественных условиях. С их помощью для расчета первичной продукции получены несколько типов уравнений, соответствующих модельным кривым (Страшкраба, Гнаук, 1989):
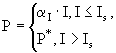 (10)
(10)
![]() , (11)
, (11)
![]() , (12)
, (12)
![]() , (13)
, (13)
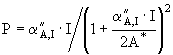 , (14)
, (14)
![]() , (15)
, (15)
где ![]() ,
, ![]() ,
, ![]() ,I - интенсивность света (ФАР),
,I - интенсивность света (ФАР), ![]() - интенсивность света, при которой достигается насыщение,
- интенсивность света, при которой достигается насыщение, ![]() - максимальная величина первичной продукции; А - фотосинтетическая способность (фотосинтетическая активность) фитопланктона, то есть количество продукции, вырабатываемой на единицу биомассы,
- максимальная величина первичной продукции; А - фотосинтетическая способность (фотосинтетическая активность) фитопланктона, то есть количество продукции, вырабатываемой на единицу биомассы, ![]() - максимальная величина фотосинтетической активности. Величина биомассы фитопланктона выражается через концентрацию хлорофилла "а". Коллекция наиболее часто используемых моделей процессов, лежащих в основе продуктивности фитопланктона, представлена Беренфельдом и Фальковским (Behrenfeld, Falkowski, 1997).
- максимальная величина фотосинтетической активности. Величина биомассы фитопланктона выражается через концентрацию хлорофилла "а". Коллекция наиболее часто используемых моделей процессов, лежащих в основе продуктивности фитопланктона, представлена Беренфельдом и Фальковским (Behrenfeld, Falkowski, 1997).
Модели делятся на два класса в зависимости от того, учитывается в них, или не учитывается фотоингибирующий фактор как реакция на заданную освещенность. В модели, широко применяемые в настоящее время в лимнологии, входит параметр ![]() , выражающий определенную (предельную) интенсивность света, при которой фотосинтез достигает максимальных значений. Величина этого параметра в разных моделях различна. В теоретическом плане наклон
, выражающий определенную (предельную) интенсивность света, при которой фотосинтез достигает максимальных значений. Величина этого параметра в разных моделях различна. В теоретическом плане наклон ![]() (или
(или ![]() ,
, ![]() ) касательной к кривой фотосинтеза, соответствующий слабой интенсивности света, определяется максимальным выходом величины хлорофилла "а" в процессе фотосинтеза. Поэтому на данном участке кривой отчетливо просматривается влияние света (Crill, 1977). Исследования показали, что величины
) касательной к кривой фотосинтеза, соответствующий слабой интенсивности света, определяется максимальным выходом величины хлорофилла "а" в процессе фотосинтеза. Поэтому на данном участке кривой отчетливо просматривается влияние света (Crill, 1977). Исследования показали, что величины ![]() ,
, ![]() ,
, ![]() зависят в основном от двух факторов: количества хлорофилла "а" в водорослях и размерного состава клеток. Содержание хлорофилла отражает световые условия существования водорослей. При этом наиболее высокое его содержание соответствует низкой освещенности I. Ферментативные процессы вступают в действие при более сильной освещенности и тогда процесс фотосинтеза начинает зависеть от температуры, а также ряда других параметров, которые выражаются через
зависят в основном от двух факторов: количества хлорофилла "а" в водорослях и размерного состава клеток. Содержание хлорофилла отражает световые условия существования водорослей. При этом наиболее высокое его содержание соответствует низкой освещенности I. Ферментативные процессы вступают в действие при более сильной освещенности и тогда процесс фотосинтеза начинает зависеть от температуры, а также ряда других параметров, которые выражаются через ![]() (Steel, 1971).
(Steel, 1971).
В тех случаях, когда специальные световые измерения невозможны, при вычислении первичной продукции можно использовать величины падающей на поверхность водоема суммарной солнечной радиации и прозрачности воды по диску Секки (Пырина, 1979). Получить их весьма несложно. Они надежны и, что особенно важно, сравнимы с аналогичными данными других авторов, поскольку основаны на стандартных методах измерения. Весь расчет строится на двух эмпирических зависимостях - проникающей суммарной солнечной радиации в глубь водоема от прозрачности воды и интенсивности фотосинтеза на разных горизонтах от энергии поступившей в них радиации.
Зависимость проникающей радиации от прозрачности воды по диску Секки, исследованная на большом материале с разнообразных водоемов, в общем виде выражается уравнением (Арэ, Толстяков, 1969):
![]() , (16)
, (16)
где ![]() - энергия солнечной радиации на глубине Z,
- энергия солнечной радиации на глубине Z, ![]() - энергия входящей в воду радиации, то есть падающей
- энергия входящей в воду радиации, то есть падающей ![]() , с поправкой на альбедо водной поверхности А:
, с поправкой на альбедо водной поверхности А: ![]() ,
, ![]() - показатель вертикального ослабления радиации, связанный с глубиной видимости диска Секки
- показатель вертикального ослабления радиации, связанный с глубиной видимости диска Секки ![]() следующим эмпирическим соотношением:
следующим эмпирическим соотношением:
![]() . (17)
. (17)
Среднесуточные величины альбедо водной поверхности разных водоемов колеблются в небольших пределах: 6 - 9%, чаще около 7%. Поэтому, если конкретных данных по альбедо нет, его принимают равным 7%. Зависимость между проникающей в воду радиацией и относительным фотосинтезом (первичной продукцией) фитопланктона, различающегося по происхождению (разные водохранилища Средней и Нижней Волги) и составу массовых видов водорослей (диатомовые и синезеленые), оказалось возможным охарактеризовать единой с точки зрения положения области насыщения световой кривой. Приближенно эта зависимость может быть представлена аналитически уравнением следующего вида:
![]() , (18)
, (18)
где ![]() и
и ![]() - соответственно относительная интенсивность фотосинтеза (первичная продукция) и энергия радиации на глубине Z. Относительная величина фотосинтеза рассчитывается в процентах к максимальному по вертикальному профилю фотосинтезу, то есть полученному при оптимальном освещении и соответствует энергии радиации на той или иной глубине. Относительные величины
- соответственно относительная интенсивность фотосинтеза (первичная продукция) и энергия радиации на глубине Z. Относительная величина фотосинтеза рассчитывается в процентах к максимальному по вертикальному профилю фотосинтезу, то есть полученному при оптимальном освещении и соответствует энергии радиации на той или иной глубине. Относительные величины ![]() затем пересчитывают в абсолютные
затем пересчитывают в абсолютные ![]() , исходя из интенсивности максимального фотосинтеза исследуемого фитопланктона
, исходя из интенсивности максимального фотосинтеза исследуемого фитопланктона ![]() :
:
![]() . (19)
. (19)
Возможно, что для других типов водоемов значения параметров уравнений придется изменять. Однако, общий вид эмпирических зависимостей останется при этом неизменным.
Введение Таллингом фотосинтетических интегралов IZ позволило перейти от кратковременных наблюдений с неизменной интенсивностью света к длительным экспериментам с меняющейся радиацией. Интегрирование ведется по глубине водоема Z, при этом верхний предел интегрирования определяется толщиной слоя перемешивания ![]() :
:
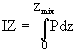 . (20)
. (20)
Изменение фотосинтеза с глубиной в данном водоеме зависит от интенсивности проникающего светового потока, ослабления света и световой чувствительности популяции. При неравномерном распределении фитопланктона по глубине ослабление света происходит по-разному, однако этот факт исследователи обычно не принимают во внимание. Суточное интегрирование для заданной глубины часто сочетается с интегралом по глубине:
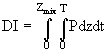 . (21)
. (21)
Существует два способа вычисления DI: 1) графическое или численное интегрирование с учетом нерегулярности суточных изменений освещенности или фотосинтеза (Fee, 1973); 2) аналитическое интегрирование в предположении синусоидального или ступенчатого суточного хода освещенности и одинакового ослабления света на всех глубинах. При этом спектральный состав света не учитывается, хотя и известно, что различные виды водорослей в соответствие со своим пигментным составом характеризуется различными "спектрами действия" (Kirck, 1975). Не принимаются во внимание и возможные изменения ![]() в течение дня как результат световой адаптации, синхронного деления популяций и недостатка питательных веществ (Harris, 1978). Кроме того, ни в одной модели не предусматривается реакция водорослей на изменение продолжительности дневного времени суток.
в течение дня как результат световой адаптации, синхронного деления популяций и недостатка питательных веществ (Harris, 1978). Кроме того, ни в одной модели не предусматривается реакция водорослей на изменение продолжительности дневного времени суток.
Использование данных моделей осложняется значительным количеством эмпирических величин, таких как ![]() , коэффициент поглощения солнечной радиации для каждой группы водорослей, фотопериод. Кроме того, в расчетные формулы входит величина толщины слоя перемешивания
, коэффициент поглощения солнечной радиации для каждой группы водорослей, фотопериод. Кроме того, в расчетные формулы входит величина толщины слоя перемешивания ![]() , нахождение которой само представляет весьма непростую физическую задачу и должно определяться в гидродинамическом блоке модели. Мы не располагаем также достаточной информацией о значении параметров для многих видов водорослей. Эти параметры не являются постоянными величинами и их трудно измерить. Модели должны явным образом учитывать физиологические процессы, а не носить характер аппроксимаций. Отчасти эти вопросы решаются в специальных моделях роста фитопланктона, с помощью которых исследуются те или иные механизмы процессов трансформации биогенных веществ в клетках.
, нахождение которой само представляет весьма непростую физическую задачу и должно определяться в гидродинамическом блоке модели. Мы не располагаем также достаточной информацией о значении параметров для многих видов водорослей. Эти параметры не являются постоянными величинами и их трудно измерить. Модели должны явным образом учитывать физиологические процессы, а не носить характер аппроксимаций. Отчасти эти вопросы решаются в специальных моделях роста фитопланктона, с помощью которых исследуются те или иные механизмы процессов трансформации биогенных веществ в клетках.
Цель моделей роста микроводорослей - предсказание скорости их роста и изменения внутриклеточного содержания углерода, азота, фосфора, концентрации хлорофилла "а" и других пигментов при заданных условиях среды, таких как температура, соленость (минерализация), доступность биогенных элементов, световые условия. Решение этих задач возможно только при соответствующем выборе структуры моделей (Zonneveld, 1997; Zonneveld, 1998). Прежде всего, в такого рода моделях в качестве переменных состояния должны рассматриваться внутриклеточные характеристики (концентрации). Во-вторых, если внутриклеточные характеристики интерпретируются как клеточные квоты, а не как отношения между биогенными элементами, то модели должны быть сформулированы на уровне отдельной клетки. В-третьих, процедура построения модели должна исходить из представлений о процессах внутри клетки и их конкретных механизмах. В качестве ключевого параметра при описании роста фитопланктона обычно рассматривается отношение хлорофилла "а" к углероду в клетке. Величина этого параметра изменяется в соответствии с изменением питательных веществ и освещенности.
Как правило, в настоящее время модели роста микроводорослей разрабатываются в результате проведения специальных лабораторных экспериментов. Культивирование водорослей в лабораторных альгоценозах возможно при различных условиях, при которых легко обеспечить определенные режимы температур, освещенности, отсутствие консументов и редуцентов, а также строгий контроль поступающих биогенных элементов. В этом состоит не только большое преимущество искусственных альгоценозов, но и главный их недостаток. С одной стороны, при изучении культур водорослей обеспечивается адекватность эмпирического материала и математической модели, учитывающей только немногие из факторов, влияющих на рост фитопланктона. Такая модель может быть тщательно откалибрована и верифицирована и, кроме того, позволяет достаточно детально изучать роль отдельных факторов, в частности исследовать закономерности смены доминирующих видов в зависимости от различных соотношений биогенных элементов. Возможность проведения необходимых экспериментов с искусственным альгоценозом позволяет проверять некоторые предположения и гипотезы, которые могут быть затем использованы в более сложных моделях реальных экосистем. С другой стороны, не следует придавать такого рода моделям и чрезмерного значения, поскольку влияние масштабов (как геометрических, так и временных) процессов, большое число факторов, действующих в реальных экосистемах и неучтенных по тем или иным причинам в искусственных альгоценозах, могут существенным образом повлиять на механизмы функционирования целостного многовидового фитопланктонного сообщества и, как следствие, на результаты моделирования.
Модели роста микроводорослей обычно строятся на основе уравнения баланса углерода в клетке. Его увеличение связано с процессом фотосинтеза, а потери обусловлены тратами на дыхание и экскрецию. Различие между поступлением и потерями определяет изменение биомассы фитопланктона. Таким образом, в общем виде уравнение динамики биомассы можно записать следующим образом:
фотосинтез - дыхание - экскреция = изменение биомассы. (22)
Очень часто при моделировании вклад экскреции не учитывается, поскольку в нормальной клетке она обычно составляет около 5% от величины связанного при фотосинтезе углерода (Fogg, 1983; Williams, 1990). Траты на дыхание параметризуют либо некоторой константой, либо считают, что они пропорциональны изменению биомассы (Sakshaug et al., 1989). Таким образом, во всех моделях основное внимание уделяется тому, каким образом описать собственно процесс фотосинтеза и его зависимость от световых условий, доступности биогенов, продолжительности и смены режимов освещения и т. п. Различия между моделями роста микроводорослей определяются, в основном, именно разными подходами к описанию фотосинтеза.
Было предложено несколько моделей роста фитопланктона, в которых описывались как процессы роста микроводорослей, так и временная изменчивость некоторых внутриклеточных структурных компонентов под влиянием различных световых условий и условий биогенного лимитирования (Bannister, 1979; Shuter, 1979; Laws, Bannister, 1980; Kiefer, Mitchell, 1983). Хотя во многих отношениях уравнения этих моделей достаточно похожи, тем не менее имеются и важные отличия в теоретических подходах, предположениях и результатах. Например, во всех упомянутых выше моделях рассчитываются величины отношений ![]() и скорости роста, однако модель (Shuter, 1979) дает возможность вычислять еще и зависимость между внутриклеточными отношениями (биогенный элемент : C) и скоростью роста. Хотя все модели являются стационарными моделями, тем не менее некоторые их этих моделей (Bannister, 1979; Laws, Bannister, 1980) способны описывать изменение интенсивности процесса фотосинтеза в ответ на кратковременные изменения освещенности. Все эти модели успешно описывают данные, полученные в лабораторных экспериментах по выращиванию микроводорослей в различных условиях. Более поздние экспериментальные работы (Falkowski et al., 1985; Goldman, 1986; Chalup, Laws, 1990) показали тем не менее, что некоторые предположения и предсказания имеющихся моделей являются ошибочными. Например, модели (Bannister, 1979; Laws, Bannister, 1980; Kiefer, Mitchell, 1983) предполагают, что среднее значение удельного коэффициента поглощения излучения хлорофиллом является постоянной величиной. Однако, исследования (Falkowski et al., 1985) показали, что значение этого коэффициента при изменении освещенности может меняться в два раза в зависимости от условий биогенного питания водорослей. Аналогичное изменение данного коэффициента наблюдается и при неизменной освещенности, но при изменении биогенного лимитирования (Chalup, Laws, 1990). Рассмотрим для примера некоторые из таких моделей.
и скорости роста, однако модель (Shuter, 1979) дает возможность вычислять еще и зависимость между внутриклеточными отношениями (биогенный элемент : C) и скоростью роста. Хотя все модели являются стационарными моделями, тем не менее некоторые их этих моделей (Bannister, 1979; Laws, Bannister, 1980) способны описывать изменение интенсивности процесса фотосинтеза в ответ на кратковременные изменения освещенности. Все эти модели успешно описывают данные, полученные в лабораторных экспериментах по выращиванию микроводорослей в различных условиях. Более поздние экспериментальные работы (Falkowski et al., 1985; Goldman, 1986; Chalup, Laws, 1990) показали тем не менее, что некоторые предположения и предсказания имеющихся моделей являются ошибочными. Например, модели (Bannister, 1979; Laws, Bannister, 1980; Kiefer, Mitchell, 1983) предполагают, что среднее значение удельного коэффициента поглощения излучения хлорофиллом является постоянной величиной. Однако, исследования (Falkowski et al., 1985) показали, что значение этого коэффициента при изменении освещенности может меняться в два раза в зависимости от условий биогенного питания водорослей. Аналогичное изменение данного коэффициента наблюдается и при неизменной освещенности, но при изменении биогенного лимитирования (Chalup, Laws, 1990). Рассмотрим для примера некоторые из таких моделей.
В модели (Laws, Chalup, 1990) предполагается, что углерод в клетке фитопланктона распределен в четырех компартментах. В соответствии с этим выделяются следующие модельные переменные: структурный углерод (S), резервный или запасенный углерод (R), углерод, связанный со световыми реакциями фотосинтеза (P) и углерод, связанный с темновыми реакциями фотосинтеза (E). Следовательно, общее количество углерода в клетке С равно сумме количеств углерода в каждом компартменте: ![]() . Отношение
. Отношение ![]() в P-компартменте обозначим через
в P-компартменте обозначим через ![]() . Предполагается, что отношение
. Предполагается, что отношение ![]() в каждом из компартментов S, P и E одинаково и равно
в каждом из компартментов S, P и E одинаково и равно ![]() . Предполагается также, что компартмент R целиком состоит из карбогидратов и липидов и, следовательно, не содержит азота. Доля углерода в структурном компартменте считается неизменной и независящей от условий роста клетки фитопланктона. В стационарном состоянии валовая скорость фотосинтеза клетки
. Предполагается также, что компартмент R целиком состоит из карбогидратов и липидов и, следовательно, не содержит азота. Доля углерода в структурном компартменте считается неизменной и независящей от условий роста клетки фитопланктона. В стационарном состоянии валовая скорость фотосинтеза клетки ![]() описывается следующим соотношением:
описывается следующим соотношением:
![]() , (23)
, (23)
где I - падающая радиация, ![]() - постоянная,
- постоянная, ![]() - некоторая функция, которая задает влияние условий среды. Скорость дыхания задается в модели следующим уравнением:
- некоторая функция, которая задает влияние условий среды. Скорость дыхания задается в модели следующим уравнением:
![]() , (24)
, (24)
где ![]() - скорость дыхания клетки в отсутствие роста,
- скорость дыхания клетки в отсутствие роста, ![]() - эмпирический коэффициент. Таким образом принимается, что скорость дыхания клетки в процессе ее роста пропорциональна валовой скорости фотосинтеза (Laws, Bannister, 1980). Предполагается, что в процессе дыхания расходуется углерод из резервного компартмента R.
- эмпирический коэффициент. Таким образом принимается, что скорость дыхания клетки в процессе ее роста пропорциональна валовой скорости фотосинтеза (Laws, Bannister, 1980). Предполагается, что в процессе дыхания расходуется углерод из резервного компартмента R.
Из вышеупомянутых предположений следует, что скорость ассимиляции питательных веществ ![]() должна удовлетворять соотношению
должна удовлетворять соотношению
![]() , (25)
, (25)
а скорость изменения углерода в клетке описывается уравнением
![]() . (26)
. (26)
В условиях сбалансированного роста удельная скорость изменения любого компонента Х задается следующим образом:
![]() . (27)
. (27)
С учетом этого уравнение (25) можно переписать так:
![]() . (28)
. (28)
Поскольку ![]() , то уравнение (26) можно преобразовать к следующему виду:
, то уравнение (26) можно преобразовать к следующему виду:
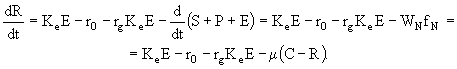 (29)
(29)
Поскольку ![]() , то, как следует из последнего уравнения,
, то, как следует из последнего уравнения,
![]() . (30)
. (30)
Из уравнения (24) получаем, что
![]() . (31)
. (31)
Из уравнения (30) с учетом соотношений для ![]() и Е получим:
и Е получим:
![]() . (32)
. (32)
Полагая, что 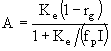 и
и ![]() , последнее уравнение можно переписать в виде квадратного уравнения относительно отношения
, последнее уравнение можно переписать в виде квадратного уравнения относительно отношения ![]() :
:
![]() . (33)
. (33)
Корни данного уравнения должны удовлетворять условию ![]() . Поэтому решение будет единственным и имеет вид:
. Поэтому решение будет единственным и имеет вид:
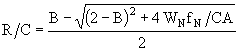 . (34)
. (34)
Оно изменяется между 0 и ![]() . Уравнение (28) можно записать в следующем виде:
. Уравнение (28) можно записать в следующем виде:
![]() . (35)
. (35)
Подставляя сюда корень квадратного уравнения, получаем:
![]() . (36)
. (36)
Значения параметров А и В зависят от внешних условий. При заданных условиях и известной величине отношения ![]() распределение углерода по выделенным в модели компартментам можно рассчитать с помощью уравнений (34), (31) и (23). Таким образом, необходимо сделать какие-то предположения относительно выбора отношения
распределение углерода по выделенным в модели компартментам можно рассчитать с помощью уравнений (34), (31) и (23). Таким образом, необходимо сделать какие-то предположения относительно выбора отношения ![]() . В модели принято, что углеродный состав клетки формируется таким образом, чтобы при данных условиях скорость ее роста была максимальной. Из уравнения (36) ясно видно, что между величиной отношения
. В модели принято, что углеродный состав клетки формируется таким образом, чтобы при данных условиях скорость ее роста была максимальной. Из уравнения (36) ясно видно, что между величиной отношения ![]() и скоростью роста
и скоростью роста ![]() существует положительная корреляция. Следовательно, величина
существует положительная корреляция. Следовательно, величина ![]() достигает максимума, когда максимально отношение
достигает максимума, когда максимально отношение ![]() . Из уравнения (34) следует, что между отношениями
. Из уравнения (34) следует, что между отношениями ![]() и
и ![]() существует отрицательная корреляция и, таким образом,
существует отрицательная корреляция и, таким образом, ![]() достигает максимума, когда
достигает максимума, когда ![]() . Если это условие выполняется, то тогда
. Если это условие выполняется, то тогда ![]() . Из уравнения (28) получаем, что в условиях биогенного насыщения
. Из уравнения (28) получаем, что в условиях биогенного насыщения
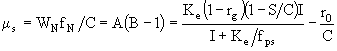 , (37)
, (37)
где ![]() ,
, ![]() - скорость роста и значение функции
- скорость роста и значение функции ![]() при насыщении. Заметим, что уравнение (37) задает гиперболическую зависимость между
при насыщении. Заметим, что уравнение (37) задает гиперболическую зависимость между ![]() и величиной освещенности I.
и величиной освещенности I.
В условиях биогенного лимитирования максимальное значение ![]() будет определяться внеклеточной концентрацией биогенов. Следовательно, в этом случае скорость роста
будет определяться внеклеточной концентрацией биогенов. Следовательно, в этом случае скорость роста ![]() будет меньше
будет меньше ![]() и
и ![]() . Из уравнений (35), (36) получим, что
. Из уравнений (35), (36) получим, что
![]() . (38)
. (38)
Заметим, что отношение ![]() клетки равно
клетки равно ![]() . С учетом этого из уравнения (38) получаем:
. С учетом этого из уравнения (38) получаем:
![]() . (39)
. (39)
В работе (Goldman, 1986) показано, что между величиной отношения ![]() и относительной скоростью роста
и относительной скоростью роста ![]() существует тесная связь, и, как показано в (Chalup, Laws, 1990), эта связь линейная. Если принять это, то в условиях насыщения, когда
существует тесная связь, и, как показано в (Chalup, Laws, 1990), эта связь линейная. Если принять это, то в условиях насыщения, когда ![]() , а
, а ![]() получаем уравнение для расчета отношения
получаем уравнение для расчета отношения ![]() :
:
![]() , (40)
, (40)
где F - весовой фактор, который меняется от 0 до 1. Используя уравнения (40), (39) и формулы для определения параметров A, B, можно получить выражение для определения функции ![]() :
:
![]() . (41)
. (41)
В работе (Falkowski et al., 1985) показано, что существует отрицательная корреляция между ![]() и I, однако конкретный вид функциональной зависимости между этими величинами установлен не был. В данном исследовании сделано предположение о том, что подходящей зависимостью может быть следующее соотношение:
и I, однако конкретный вид функциональной зависимости между этими величинами установлен не был. В данном исследовании сделано предположение о том, что подходящей зависимостью может быть следующее соотношение:
![]() , (42)
, (42)
где ![]() - значение
- значение ![]() при
при ![]() , а
, а ![]() - это значение I, когда
- это значение I, когда ![]() . Подстановка (42) в (37) дает возможность определить величину
. Подстановка (42) в (37) дает возможность определить величину ![]() :
:
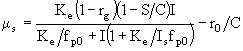 . (43)
. (43)
Из уравнений (38), (31) и соотношений для определения параметров А и В можно получить выражение для вычисления отношения E/C:
![]() . (44)
. (44)
С его помощью легко найти выражения для расчета отношения ![]() :
:
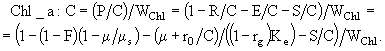 (45)
(45)
Наконец, с помощью уравнений (24) и (44) можно найти отношение
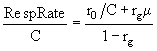 , (46)
, (46)
которое дает возможность определить затраты на дыхание водорослей.
Клеточная квота углерода многих видов фитопланктона изменяется при изменении интенсивности освещенности (изменении плотности фотонов). Обычно, она увеличивается с ростом интенсивности падающего излучения (Laws, Bannister, 1980; Falkowski et al., 1985). Однако модели, основанные на балансовом равенстве (22), часто не учитывают этого. В работе (Zonneveld et al., 1997) предлагается модель, которая рассматривает возможный механизм изменения клеточной квоты углерода и удельной скорости роста в зависимости от изменений интенсивности потока освещенности. Принимается, что весь запас углерода в клетке распределен всего в двух компартментах (пулах): переходном (или временном) пуле C и пуле постоянного объема V. При этом выделяют три процесса, в результате которых происходит изменение углерода в клетке: процесс ассимиляции (A), в результате которого увеличивается переходный пул углерода, клеточный метаболизм (M) и рост (G). Все три процесса зависят от внешних условий, в частности, от условий освещения, температуры и концентрации ![]() . Однако, в данной модели, ради ее упрощения, принято, что внешние факторы (свет и
. Однако, в данной модели, ради ее упрощения, принято, что внешние факторы (свет и ![]() ) влияют только на процесс ассимиляции клеткой углерода. Размер клетки, или ее биомасса, определяется суммой углерода в двух выделенных пулах. Под ростом понимается увеличение биомассы пула постоянного объема. Деление клетки происходит в том и только в том случае, когда биомасса углерода в этом компартменте удвоится. При увеличении интенсивности потока освещенности клетка может ассимилировать большее количество углерода в переходном компартменте. Одновременно с этим постепенно будет расти и масса углерода в пуле постоянного объема, хотя изменение освещенности напрямую не влияет на содержание в нем углерода. В модели принимается, что ассимиляция и внутренний метаболизм зависят от содержания углерода в компартменте V, а процесс роста - от отношения
) влияют только на процесс ассимиляции клеткой углерода. Размер клетки, или ее биомасса, определяется суммой углерода в двух выделенных пулах. Под ростом понимается увеличение биомассы пула постоянного объема. Деление клетки происходит в том и только в том случае, когда биомасса углерода в этом компартменте удвоится. При увеличении интенсивности потока освещенности клетка может ассимилировать большее количество углерода в переходном компартменте. Одновременно с этим постепенно будет расти и масса углерода в пуле постоянного объема, хотя изменение освещенности напрямую не влияет на содержание в нем углерода. В модели принимается, что ассимиляция и внутренний метаболизм зависят от содержания углерода в компартменте V, а процесс роста - от отношения ![]() .
.
Для описания процесса ассимиляции в данной модели используется гиперболическая зависимость между ассимиляцией и плотностью потока освещенности I:
![]() , (47)
, (47)
где ![]() - величина максимальной ассимиляции на единицу пула постоянного объема,
- величина максимальной ассимиляции на единицу пула постоянного объема, ![]() - плотность потока освещенности, при котором ассимиляция равна половине от максимального ее значения при данных условиях. В модели не принимается во внимание влияние освещенности на величину
- плотность потока освещенности, при котором ассимиляция равна половине от максимального ее значения при данных условиях. В модели не принимается во внимание влияние освещенности на величину ![]() . Для описания клеточного метаболизма используется простейшая, линейная, его зависимость от переменной V (реакция первого порядка):
. Для описания клеточного метаболизма используется простейшая, линейная, его зависимость от переменной V (реакция первого порядка):
![]() . (48)
. (48)
В соответствие с рассматриваемым в модели механизмом роста, этот процесс описывается следующим уравнением:
![]() . (49)
. (49)
Динамика углерода в переходном компартменте описывается простым кинетическим уравнением реакции первого порядка:
![]() , (50)
, (50)
где ![]() - эмпирическая константа. Физиологическая интерпретация этого параметра состоит в том, что в условиях светового насыщения, когда
- эмпирическая константа. Физиологическая интерпретация этого параметра состоит в том, что в условиях светового насыщения, когда ![]() и количество углерода в переходном пуле достигает предельной для данных условий величины
и количество углерода в переходном пуле достигает предельной для данных условий величины ![]() , его величина будет равна отношению
, его величина будет равна отношению ![]() . Таким образом, с помощью уравнений (48) - (50) можно описать скорость изменения углерода, связанного с процессами метаболизма и роста клетки. Удельная скорость роста клетки
. Таким образом, с помощью уравнений (48) - (50) можно описать скорость изменения углерода, связанного с процессами метаболизма и роста клетки. Удельная скорость роста клетки ![]() , как показано в данной работе, представляет собой функцию содержания углерода в переходном компартменте и определяется следующим соотношением:
, как показано в данной работе, представляет собой функцию содержания углерода в переходном компартменте и определяется следующим соотношением:
![]() . (51)
. (51)
В стационарном состоянии, когда ![]() , удельная скорость роста
, удельная скорость роста ![]() , а стационарная концентрация углерода в переменном компартменте -
, а стационарная концентрация углерода в переменном компартменте -
![]() . (52)
. (52)
Подставляя (52) в (51), получим соотношение для определения стационарной скорости роста ![]() как функцию интенсивности освещенности:
как функцию интенсивности освещенности:
![]() . (53)
. (53)
Преобразуем последнее соотношение. Обозначим через ![]() максимальную скорость роста,
максимальную скорость роста, ![]() ,
, ![]() . Тогда из (53) получим:
. Тогда из (53) получим:
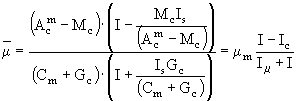 . (54)
. (54)
Таким образом, полученные уравнения (52) - (54) определяют искомую зависимость клеточной квоты углерода и удельной скорости роста в зависимости от изменений интенсивности потока освещенности. Заметим, что в данной модели отношение ![]() в клетке не играет никакой роли и этим данная модель отличается от многих других моделей аналогичного типа, в которых это отношение является входной переменной (Laws, Bannister, 1980; Kiefer, Mitchell, 1983; Sakshaug et al., 1989). Это вовсе не означает, что модели, использующие величины хлорофилла "а" в качестве меры биомассы фитопланктона хуже рассмотренной модели. Все зависит от конкретной постановки задачи. В данной работе исследовался механизм влияния интенсивности освещенности на клеточную квоту углерода. Однако для практической океанологии или лимнологии, именно использование отношения
в клетке не играет никакой роли и этим данная модель отличается от многих других моделей аналогичного типа, в которых это отношение является входной переменной (Laws, Bannister, 1980; Kiefer, Mitchell, 1983; Sakshaug et al., 1989). Это вовсе не означает, что модели, использующие величины хлорофилла "а" в качестве меры биомассы фитопланктона хуже рассмотренной модели. Все зависит от конкретной постановки задачи. В данной работе исследовался механизм влияния интенсивности освещенности на клеточную квоту углерода. Однако для практической океанологии или лимнологии, именно использование отношения ![]() оказывается более предпочтительным, поскольку концентрация хлорофилла легко и достаточно точно измеряется даже со спутников.
оказывается более предпочтительным, поскольку концентрация хлорофилла легко и достаточно точно измеряется даже со спутников.
В работе (Zonneveld, 1998) проведено исследование, цель которого состояла в том, чтобы предсказать, как будет меняться отношение хлорофилл "а" : C при изменении условий среды. Для этого рассматривается клеточная модель, которая позволяет описывать клеточные квоты углерода и хлорофилла "а" и их временную динамику. В данной модели клетка фитопланктона характеризуется тремя переменными состояния: постоянным объемом (массой) с фиксированным отношением C:N и двумя переменными пулами питательных веществ - один для углерода, другой - для азота. Процесс ассимиляции приводит к увеличению переменных пулов питательных веществ. Принята во внимание зависимость ассимиляции углерода от условий освещенности. Возможное влияние освещенности на ассимиляцию азота в данной модели не рассматривается. Углерод и азот мобилизуются из пулов для того, чтобы поддерживать постоянную массу. Изменения в величине углеродного и азотного пулов по отношению к постоянной массе приводят к изменению элементного состава клетки.
Рост, лимитированный питательными веществами. Пусть N обозначает количество азота в переменном азотном пуле. Часто оказывается более удобным работать не с концентрацией, а с плотностью азота ![]() : количеством азота в пуле, деленном на постоянный объем V. Это исключает необходимость рассматривать в качестве переменной величины объем, занимаемый переменным пулом. Предполагается, что если клетки фитопланктона растут при насыщающих концентрациях азота в окружающей среде в течение достаточно длительного промежутка времени, то плотность азота постепенно достигает некоторого постоянного максимального значения
: количеством азота в пуле, деленном на постоянный объем V. Это исключает необходимость рассматривать в качестве переменной величины объем, занимаемый переменным пулом. Предполагается, что если клетки фитопланктона растут при насыщающих концентрациях азота в окружающей среде в течение достаточно длительного промежутка времени, то плотность азота постепенно достигает некоторого постоянного максимального значения ![]() . Это значение не может быть превышено до тех пор, пока именно питательные вещества являются лимитирующими. Предполагается также, что скорость поглощения питательных веществ пропорциональна объему клетки и зависит от концентрации биогенов в окружающей среде, а кинетика поглощения соответствует кинетике Михаэлиса-Ментен. Обозначим через
. Это значение не может быть превышено до тех пор, пока именно питательные вещества являются лимитирующими. Предполагается также, что скорость поглощения питательных веществ пропорциональна объему клетки и зависит от концентрации биогенов в окружающей среде, а кинетика поглощения соответствует кинетике Михаэлиса-Ментен. Обозначим через ![]() гиперболическую зависимость скорости поглощения от концентрации субстрата
гиперболическую зависимость скорости поглощения от концентрации субстрата ![]() , то есть
, то есть ![]() , где
, где ![]() - константа полунасыщения. Предполагается, что скорость изменения плотности азота описывается уравнением
- константа полунасыщения. Предполагается, что скорость изменения плотности азота описывается уравнением
![]() , (55)
, (55)
то есть кинетика плотности азота соответствует кинетике первого порядка. Слагаемое ![]() представляет собой использование азота из переменного пула, а
представляет собой использование азота из переменного пула, а ![]() - константа пропорциональности.
- константа пропорциональности. ![]() - максимальная скорость ассимиляции. Если внешняя концентрация азота достигает насыщения
- максимальная скорость ассимиляции. Если внешняя концентрация азота достигает насыщения ![]() и остается таковой в течение достаточно длительного периода времени, то плотность азота больше не меняется, так что
и остается таковой в течение достаточно длительного периода времени, то плотность азота больше не меняется, так что ![]() . При этих условиях получаем, что
. При этих условиях получаем, что ![]() . С одной стороны, скорость утилизации азота равна скорости его ассимиляции
. С одной стороны, скорость утилизации азота равна скорости его ассимиляции ![]() минус скорость изменения количества питательного вещества
минус скорость изменения количества питательного вещества ![]() . С другой стороны, если азот используется только для роста, без вытеснения его из клетки, и содержание азота в единице структурного (неизменного) объема
. С другой стороны, если азот используется только для роста, без вытеснения его из клетки, и содержание азота в единице структурного (неизменного) объема ![]() постоянно, то скорость его утилизации должна быть равна
постоянно, то скорость его утилизации должна быть равна ![]() , так что
, так что ![]() . Это уравнение можно разрешить относительно
. Это уравнение можно разрешить относительно ![]() - скорости роста. Переходя к удельной скорости роста
- скорости роста. Переходя к удельной скорости роста ![]() , получим:
, получим:
![]() . (56)
. (56)
Если рассматривается, например, хемостат в стационарном состоянии, то удельная скорость роста равна скорости разбавления D. Из уравнения (56) получаем, что в этом случае
![]() . (57)
. (57)
Углерод и хлорофилл "а" в фитопланктоне в условиях азотного лимитирования. Предполагается, что возможны три базовые варианта зависимости скорости поглощения от плотности лимитирующего питательного вещества: а) скорость поглощения не зависит от плотности вещества; б) скорость поглощения может быть пропорциональна плотности вещества; в) скорость поглощения может быть пропорциональна удельной скорости роста, то есть описываться гиперболической зависимостью от плотности биогенного элемента. Для конкретного вида фитопланктона можно также использовать любую линейную комбинацию из этих трех базовых вариантов. Углерод в клетке используется для метаболизма, роста и дыхания. Принимается, что метаболические потребности пропорциональны постоянному объему, дыхание - пропорционально скорости роста. Тогда скорость изменения пула углерода задается следующим уравнением:
![]() . (58)
. (58)
Скорость ассимиляции углерода задается соотношением
![]() , (59)
, (59)
где ![]() ,
, ![]() ,
, ![]() - весовые коэффициенты, соответствующие вариантам а) - в) соответственно. Предполагается, что в условиях азотного лимитирования клеток фитопланктона абсолютная скорость образования хлорофилла "а" зависит от количества азота в переменном пуле и описывается реакцией первого порядка:
- весовые коэффициенты, соответствующие вариантам а) - в) соответственно. Предполагается, что в условиях азотного лимитирования клеток фитопланктона абсолютная скорость образования хлорофилла "а" зависит от количества азота в переменном пуле и описывается реакцией первого порядка: ![]() , где k - константа. Скорость образования хлорофилла может зависеть и от условий освещенности. В этом случае k будет зависеть от интенсивности освещения. Кроме того, клеточная квота хлорофилла уменьшается вследствие роста клеток и деструкции хлорофилла, что характеризуется константой скорости деструкции r. При этих предположениях внутриклеточное изменение концентрации хлорофилла соответствует уравнению:
, где k - константа. Скорость образования хлорофилла может зависеть и от условий освещенности. В этом случае k будет зависеть от интенсивности освещения. Кроме того, клеточная квота хлорофилла уменьшается вследствие роста клеток и деструкции хлорофилла, что характеризуется константой скорости деструкции r. При этих предположениях внутриклеточное изменение концентрации хлорофилла соответствует уравнению:
![]() . (60)
. (60)
Рост, лимитированный световыми условиями. В этом случае скорость роста определяется переменным пулом углерода, а не азота. Это означает, что для описания изменчивости отношения хлорофилл "а" : С вместо уравнения, задающего скорость изменения азота в клетке, нужно ввести уравнение, задающее скорость изменения углерода. Это уравнение имеет следующий вид:
![]() . (61)
. (61)
Здесь I - интенсивность света (ФАР), ![]() - интенсивность света, при которой достигается насыщение. Углерод удовлетворяет энергетические потребности клетки и служит одним из биогенных элементов, необходимых ей для роста. Что касается энергетических потребностей, то углерод используется для того, чтобы покрыть траты на метаболизм и дыхание. Кроме того, он используется в качестве строительного материала. Таким образом, скорость утилизации углерода задается слагаемыми
- интенсивность света, при которой достигается насыщение. Углерод удовлетворяет энергетические потребности клетки и служит одним из биогенных элементов, необходимых ей для роста. Что касается энергетических потребностей, то углерод используется для того, чтобы покрыть траты на метаболизм и дыхание. Кроме того, он используется в качестве строительного материала. Таким образом, скорость утилизации углерода задается слагаемыми ![]() . С другой стороны, скорость его утилизации равна
. С другой стороны, скорость его утилизации равна ![]() . С помощью этих двух соотношений можно получить выражение для удельной скорости роста при световом лимитировании:
. С помощью этих двух соотношений можно получить выражение для удельной скорости роста при световом лимитировании:
![]() . (62)
. (62)
В работе (Krause-Jensen, Sand-Jensen, 1998) проведено исследование, которое преследовало следующие две основные цели: 1) проверить возможность существования общей зависимости между концентрацией хлорофилла а (![]() ), поглощением света и валовым фотосинтезом для любого водного растительного сообщества; 2) оценить связь между максимумом биомассы и валовым фотосинтезом различных водных растительных сообществ. Анализ был проведен на основе 414 заимствованных из литературы данных по фитопланктону, макрофитам и бентосным микроводорослям, главным образом для водоемов умеренной зоны.
), поглощением света и валовым фотосинтезом для любого водного растительного сообщества; 2) оценить связь между максимумом биомассы и валовым фотосинтезом различных водных растительных сообществ. Анализ был проведен на основе 414 заимствованных из литературы данных по фитопланктону, макрофитам и бентосным микроводорослям, главным образом для водоемов умеренной зоны.
·Поглощение света в водных растительных сообществах. В оптически однородных водных растительных сообществах монохроматическая освещенность затухает с глубиной по экспоненциальному закону в соответствии с уравнением
![]() . (63)
. (63)
Способность растительного сообщества ослаблять поток солнечной радиации в диапазоне ФАР (400 - 700 нм) удобно характеризовать величиной среднего коэффициента ослабления ![]() , а не совокупностью аналогичных коэффициентов для различных длин волн. Средний коэффициент ослабления
, а не совокупностью аналогичных коэффициентов для различных длин волн. Средний коэффициент ослабления ![]() представляет собой величину, в которой учитывается процесс ослабления собственно растениями
представляет собой величину, в которой учитывается процесс ослабления собственно растениями ![]() и фоновое ослабление
и фоновое ослабление ![]() . Ослабление, связанное с растениями, рассчитывается через концентрацию хлорофилла С и удельный коэффициент ослабления хлорофилла
. Ослабление, связанное с растениями, рассчитывается через концентрацию хлорофилла С и удельный коэффициент ослабления хлорофилла ![]() . В таком случае получаем, что
. В таком случае получаем, что
![]() . (63)
. (63)
![]() измеряется в м-1, С - в (мг Chl) м-3,
измеряется в м-1, С - в (мг Chl) м-3, ![]() - в м2 (мг Chl)-1. Аналогичное соотношение можно получить и для случая, когда концентрация выражается в единицах сухого веса. Вместо величины
- в м2 (мг Chl)-1. Аналогичное соотношение можно получить и для случая, когда концентрация выражается в единицах сухого веса. Вместо величины ![]() иногда удобнее использовать глубину фотического слоя (фотической зоны)
иногда удобнее использовать глубину фотического слоя (фотической зоны) ![]() , то есть такую глубину, на которой ослабляется 99% освещенности.
, то есть такую глубину, на которой ослабляется 99% освещенности. ![]() .
.
·Объемный фотосинтез. Максимальная величина объемного фотосинтеза ![]() (ммоль O2 м-3 ч-1) представляет собой произведение концентрации хлорофилла С (мг Chl) м-3 на максимальную величину удельного фотосинтеза
(ммоль O2 м-3 ч-1) представляет собой произведение концентрации хлорофилла С (мг Chl) м-3 на максимальную величину удельного фотосинтеза ![]() (ммоль О2 (мг Chl)-1 ч-1:
(ммоль О2 (мг Chl)-1 ч-1:
![]() . (64)
. (64)
Выражая С из (63) и подставляя в (64), получим:
![]() . (65)
. (65)
В плотном растительном сообществе ослабление света осуществляется в основном растениями и второе слагаемое в (65) мало. Поэтому
![]() . (66)
. (66)
Отсюда следует, что если отношение между максимумом удельного фотосинтеза и удельным коэффициентом ослабления в плотном сообществе примерно постоянно, то величина объемного фотосинтеза прямо пропорциональна величине среднего коэффициента ослабления ![]() и обратно пропорциональна толщине фотического слоя
и обратно пропорциональна толщине фотического слоя ![]() . Аналогичные соотношение можно получить и для случая, когда концентрация выражается в единицах сухого веса.
. Аналогичные соотношение можно получить и для случая, когда концентрация выражается в единицах сухого веса.
·Интегральный фотосинтез. Толлинг установил зависимость между величиной интегрального фотосинтеза ![]() (ммоль O2 м-2 ч-1), концентрацией хлорофилла С (мг Chl) м-3, средним коэффициентом ослабления
(ммоль O2 м-2 ч-1), концентрацией хлорофилла С (мг Chl) м-3, средним коэффициентом ослабления ![]() (м-1), падающим потоком фотонов
(м-1), падающим потоком фотонов ![]() (мкмоль фотонов м-2 с-1) и потоком, при котором наступает световое насыщение фотосинтеза
(мкмоль фотонов м-2 с-1) и потоком, при котором наступает световое насыщение фотосинтеза ![]() :
:
![]() . (67)
. (67)
Первый множитель представляет собой величину максимального объемного фотосинтеза, второй - определяет глубину, на которой ![]() в точности будет равен
в точности будет равен ![]() . Эта формула была проверена на целом ряде пресноводных экосистем. Аналогичная формула была получена и для морских условий. Для плотных сообществ растений
. Эта формула была проверена на целом ряде пресноводных экосистем. Аналогичная формула была получена и для морских условий. Для плотных сообществ растений
![]() . (68)
. (68)
Значения ![]() достаточно изменчивы, однако систематических отличий среди различных типов водных сообществ не обнаружено. С помощью формул (67) и (68) можно выразить величину интегрального фотосинтеза
достаточно изменчивы, однако систематических отличий среди различных типов водных сообществ не обнаружено. С помощью формул (67) и (68) можно выразить величину интегрального фотосинтеза ![]() по отношению к максимальному интегральному фотосинтезу
по отношению к максимальному интегральному фотосинтезу ![]() , который достигается в том случае, если весь доступный свет будет использован растительным сообществом:
, который достигается в том случае, если весь доступный свет будет использован растительным сообществом:
![]() . (69)
. (69)
Из этого выражения следует, что фоновое ослабление света выступает в качестве конкурентного ингибитора ![]() . Эта формула обусловливает тот же тип кинетики, что и в случае односубстратной ферментативной реакции. Отношение
. Эта формула обусловливает тот же тип кинетики, что и в случае односубстратной ферментативной реакции. Отношение ![]() представляет собой такую концентрацию хлорофилла, при которой достигается половина верхнего предела величины интегрального фотосинтеза.
представляет собой такую концентрацию хлорофилла, при которой достигается половина верхнего предела величины интегрального фотосинтеза.
В работе (Левич А.П. и др., 1997) разработана модель для исследования влияния различных концентраций азота и фосфора в среде на структуру лабораторного сообщества фитопланктона. Модель построена для альгоценоза, функционирующего в условиях накопительного культивирования. Процессы роста и поглощения биогенных элементов описываются раздельно. Скорость роста биомассы фитопланктона описывается гиперболической зависимостью и, согласно современным представлениям (Йоргенсен, 1985), зависит от внутренних концентраций биогенных элементов. Скорость поглощения каждого биогенного элемента из среды в рассматриваемых условиях предполагается равной скорости изменения общего внутриклеточного запаса данного элемента и определяется как его внутриклеточным, так и внешним уровнем. Помимо динамики биомассы фитопланктона и внутриклеточного содержания биогенных элементов (азота и фосфора), моделируется и динамика концентрации азота и фосфора в среде культивирования. В данном случае это сделать очень просто на основании балансовых соотношений. В соответствии с законом сохранения вещества, применительно к конкретным условиям, скорость изменения концентрации биогенного элемента в среде по абсолютной величине равна скорости поглощения этого элемента фитопланктоном.
Таким образом, для каждого вида в модель включено три уравнения: динамики биомассы и динамики внутриклеточного содержания азота и фосфора, а для всего сообщества - уравнения динамики концентраций азота и фосфора в среде. Модель записывается в виде системы дифференциальных уравнений и имеет следующий вид:
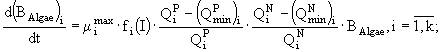 (70)
(70)
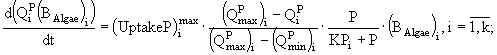 (71)
(71)
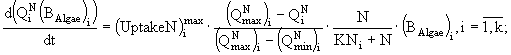 (71)
(71)
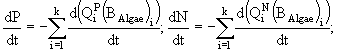 (72)
(72)
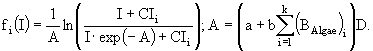 (73)
(73)
Переменные состояния модели: ![]() - концентрация биомассы i-го вида водорослей;
- концентрация биомассы i-го вида водорослей; ![]() ,
, ![]() - клеточные квоты в единице биомассы по фосфору и азоту соответственно; N и P - концентрации азота и фосфора в среде. Параметры модели:
- клеточные квоты в единице биомассы по фосфору и азоту соответственно; N и P - концентрации азота и фосфора в среде. Параметры модели: ![]() - максимальная удельная скорость роста i-го вида водорослей;
- максимальная удельная скорость роста i-го вида водорослей; ![]() ,
, ![]() - максимальные удельные скорости поглощения фосфора и азота i-м видом;
- максимальные удельные скорости поглощения фосфора и азота i-м видом; ![]() ,
, ![]() - максимальные и минимальные клеточные квоты по фосфору и азоту соответственно;
- максимальные и минимальные клеточные квоты по фосфору и азоту соответственно; ![]() ,
, ![]() - константы полунасыщения по фосфору и азоту для i-го вида фитопланктона;
- константы полунасыщения по фосфору и азоту для i-го вида фитопланктона; ![]() - константа радиации для i-го вида; I - уровень освещенности; a - коэффициент поглощения света водой; b - коэффициент поглощения света фитопланктоном; D - глубина; k - количество видов.
- константа радиации для i-го вида; I - уровень освещенности; a - коэффициент поглощения света водой; b - коэффициент поглощения света фитопланктоном; D - глубина; k - количество видов.
В данной модели учтена конкуренция за биогенные элементы и конкурентное взаимодействие путем затенения. Путем некоторого усложнения модели можно учесть и другие типы взаимодействия видов, например, посредством метаболитов, которые, по всей видимости, играют немаловажную роль в сообществах фитопланктона (Алексеев, 1976; Страшкраба, Гнаук, 1989). Возможный путь такого учета несубстратных взаимодействий видов в данной модели - предусмотреть явную зависимость физиологических параметров от состава фитопланктонного сообщества. Однако, при плохой изученности несубстратных взаимодействий видов, при отсутствии приемлемых способов их математического описания усложнение модели представляется авторам нецелесообразным.
В работе (Arrigo, Sullivan, 1994) рассматривается модель роста фитопланктона, в которой основное внимание сосредоточено на описании влияния температуры, внутрисуточных изменений спектральных характеристик освещенности, биогенных элементов (нитратного, аммонийного азота, фосфатов и силикатов), а также солености воды. Величина скорости роста микроводорослей ![]() на определенной глубине моря z определяется в модели через максимальную скорость роста, которая корректируется определенным образом на температурные, световые условия, условия биогенного питания, различную соленость воды (Arrigo, Sullivan, 1992). Максимальная удельная скорость роста фитопланктона как функция температуры воды задается в модели следующим соотношением (Eppley, 1972):
на определенной глубине моря z определяется в модели через максимальную скорость роста, которая корректируется определенным образом на температурные, световые условия, условия биогенного питания, различную соленость воды (Arrigo, Sullivan, 1992). Максимальная удельная скорость роста фитопланктона как функция температуры воды задается в модели следующим соотношением (Eppley, 1972):
![]() , (74)
, (74)
где ![]() - удельная скорость роста при 0°С,
- удельная скорость роста при 0°С, ![]() - константа, которая определяет "чувствительность" максимальной скорости роста к изменениям температуры воды,
- константа, которая определяет "чувствительность" максимальной скорости роста к изменениям температуры воды, ![]() - температура воды на глубине z. Для того, чтобы учесть зависимость удельной скорости роста от других факторов, например, концентрации биогенных веществ, используется формула Моно:
- температура воды на глубине z. Для того, чтобы учесть зависимость удельной скорости роста от других факторов, например, концентрации биогенных веществ, используется формула Моно:
![]() , (75)
, (75)
где ![]() - удельная скорость роста,
- удельная скорость роста, ![]() - концентрация i-го биогенного элемента (то есть, нитратного, аммонийного азота, фосфатов и силикатов) на глубине z,
- концентрация i-го биогенного элемента (то есть, нитратного, аммонийного азота, фосфатов и силикатов) на глубине z, ![]() - константа полунасыщения для
- константа полунасыщения для ![]() , определяемая как концентрация биогенных элементов, при которой
, определяемая как концентрация биогенных элементов, при которой ![]() . Предполагается, что величина
. Предполагается, что величина ![]() не зависит от температуры. Из уравнения (75) можно получить выражение для безразмерной удельной скорости роста и ее зависимость от i-го биогенного элемента:
не зависит от температуры. Из уравнения (75) можно получить выражение для безразмерной удельной скорости роста и ее зависимость от i-го биогенного элемента:
![]() . (76)
. (76)
Уравнение (76) используется в дальнейшем для определения относительного коэффициента биогенного лимитирования ![]() на глубине z. Эта величина оценивается по минимуму безразмерной удельной скорости:
на глубине z. Эта величина оценивается по минимуму безразмерной удельной скорости:
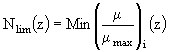 . (77)
. (77)
Многие модели описывают световое и биогенное лимитирование роста фитопланктона раздельно (Shuter, 1979; Laws, Bannister, 1980; Kiefer, Mitchell, 1983). Аналогичный подход использован и в данной работе. При этом способ описания процесса светового лимитирования имеет следующие особенности: 1) он зависит от единственного фотосинтетического параметра; 2) он учитывает спектральные изменения плотности освещенности на глубине z, которые связаны с процессами поглощения водой и пигментами фитопланктона; 3) он учитывает внутрисуточные изменения спектрального состава освещенности. Для описания функциональной зависимости между скоростью фотосинтеза и падающей радиацией используются разные соотношения. В большинстве случаев фотосинтетически активная радиация (PAR) рассчитывается с помощью следующего соотношения:
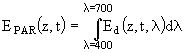 , (78)
, (78)
где ![]() - проникающая радиация на глубине z в момент времени t и с длиной волны
- проникающая радиация на глубине z в момент времени t и с длиной волны ![]() . В рассматриваемой модели зависимость между скоростью фотосинтеза на глубине z и величиной PAR на этой же глубине описывается следующим уравнением (Platt et al., 1980):
. В рассматриваемой модели зависимость между скоростью фотосинтеза на глубине z и величиной PAR на этой же глубине описывается следующим уравнением (Platt et al., 1980):
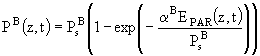 , (79.1)
, (79.1)
где ![]() - мгновенная скорость фотосинтеза на глубине z в момент времени t;
- мгновенная скорость фотосинтеза на глубине z в момент времени t; ![]() - предельная при данной освещенности скорость фотосинтеза;
- предельная при данной освещенности скорость фотосинтеза; ![]() - эффективность фотосинтеза, которая определяется как тангенс угла наклона касательной к
- эффективность фотосинтеза, которая определяется как тангенс угла наклона касательной к ![]() -
-![]() кривой при
кривой при ![]() .
.
Уравнение (79.1) можно переписать, используя для этого параметр фотоадаптации ![]() (
(![]() ) в отсутствии фотоингибирования:
) в отсутствии фотоингибирования:
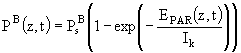 . (79.2)
. (79.2)
Предполагается, что в течение суток параметр ![]() остается неизменным. Для того, чтобы получить функцию светового лимитирования в безразмерной форме, в модели вводится корректирующий множитель
остается неизменным. Для того, чтобы получить функцию светового лимитирования в безразмерной форме, в модели вводится корректирующий множитель ![]() , который представляет собой следующее отношение:
, который представляет собой следующее отношение:
![]() .
.
Тогда функцию светового лимитирования в безразмерном (нормированном) виде можно записать следующим образом:
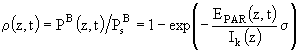 . (80)
. (80)
Принято, что внутриклеточное отношение ![]() зависит от фотопериода D и величины PAR и в общем виде эту зависимость можно представить так:
зависит от фотопериода D и величины PAR и в общем виде эту зависимость можно представить так:
![]() , (81)
, (81)
где a, b - некоторые эмпирические коэффициенты.
Существенным недостатком формулы (80) является то, что она не учитывает изменение спектрального состава света. Для того, чтобы ввести зависимость освещенности от длины волны, вместо величины PAR рассматривается величина фотосинтетически используемой солнечной радиации (PUR) (Morel, 1978), которая представляет собой такую долю от ФАР, которая реально используется при фотосинтезе:
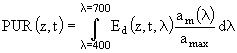 . (82)
. (82)
Здесь ![]() - спектр поглощения для данной пробы микроводорослей,
- спектр поглощения для данной пробы микроводорослей, ![]() - максимум поглощения в диапазоне длин волн от 400 до 700 нм. С учетом этого уравнение (80) перепишется в следующем виде:
- максимум поглощения в диапазоне длин волн от 400 до 700 нм. С учетом этого уравнение (80) перепишется в следующем виде:
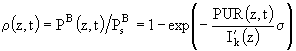 , (83)
, (83)
где ![]() имеет тот же самый смысл, что и
имеет тот же самый смысл, что и ![]() . Внутрисуточная изменчивость
. Внутрисуточная изменчивость ![]() описывается как синусоидальная функция от времени t на промежутке от
описывается как синусоидальная функция от времени t на промежутке от ![]() до
до ![]() , где D - величина фотопериода для данного времени года. Таким образом, получаем:
, где D - величина фотопериода для данного времени года. Таким образом, получаем:
![]() . (84)
. (84)
В данной модели величина ![]() , ради упрощения, описывается в виде логистической зависимости от среднесуточной величины PUR. Общий вид зависимости получен на основании многочисленных данных наблюдений. Ее можно записать в следующем виде:
, ради упрощения, описывается в виде логистической зависимости от среднесуточной величины PUR. Общий вид зависимости получен на основании многочисленных данных наблюдений. Ее можно записать в следующем виде:
![]() , (85)
, (85)
где A и B - эмпирические параметры.
Для того, чтобы учесть влияние нескольких факторов на скорость роста фитопланктона, в данной модели использована следующая мультипликативная зависимость:
![]() , (86)
, (86)
где ![]() ,
, ![]() - среднесуточный коэффициент светового лимитирования,
- среднесуточный коэффициент светового лимитирования, ![]() - (в данной работе) полиномиальная функция, описывающая влияние солености на удельную скорость роста фитопланктона. Это уравнение представляет собой одну из немногих попыток рассмотрения совместного воздействия сразу нескольких факторов на рост фитопланктона: температуры, солености, биогенного лимитирования и суточных изменений в спектральном составе освещенности.
- (в данной работе) полиномиальная функция, описывающая влияние солености на удельную скорость роста фитопланктона. Это уравнение представляет собой одну из немногих попыток рассмотрения совместного воздействия сразу нескольких факторов на рост фитопланктона: температуры, солености, биогенного лимитирования и суточных изменений в спектральном составе освещенности.
Зависимость удельной скорости дыхания микроводорослей от температуры воды ![]() на заданном горизонте z задавалась в модели следующим соотношением:
на заданном горизонте z задавалась в модели следующим соотношением:
![]() . (87)
. (87)
Здесь ![]() - удельная скорость дыхания при 0°С,
- удельная скорость дыхания при 0°С, ![]() - эмпирический коэффициент. Выедание фитопланктона описывается в модели очень просто. Чтобы не вводить еще одно уравнение для динамики нехищного зоопланктона, полагали, что удельная скорость выедания микроводорослей
- эмпирический коэффициент. Выедание фитопланктона описывается в модели очень просто. Чтобы не вводить еще одно уравнение для динамики нехищного зоопланктона, полагали, что удельная скорость выедания микроводорослей ![]() составляет некоторую часть от чистого фотосинтеза, то есть
составляет некоторую часть от чистого фотосинтеза, то есть
![]() . (88)
. (88)
Аналогично полагали, что удельная скорость экскреции ![]() биогенных элементов организмами зоопланктона составляет определенную долю от удельной скорости выедания ими фитопланктона:
биогенных элементов организмами зоопланктона составляет определенную долю от удельной скорости выедания ими фитопланктона:
![]() , (89)
, (89)
где e - эмпирический коэффициент.
Временная динамика фитопланктона и биогенных элементов на заданном горизонте z задается в виде системы балансовых дифференциальных уравнений. Мы не будем подробно останавливаться на математическом описании динамики гидрооптических компонентов модели. Эти вопросы будут рассмотрены ниже. Для нас было важно рассмотреть то, каким образом решается задача модельного описания роста фитопланктона при комбинированном воздействии внешних факторов.
Подавляющее большинство моделей водных экосистем не учитывает структуры подводных оптических полей. Применяемое рядом исследователей механическое использование моделей, развитых для одних водных объектов при исследовании других, как правило, неправомочно из-за недоучета специфических оптических свойств воды (Семовский, 1999). Дело здесь в том, что оптические свойства вод определяют процесс фотосинтетической первичной продукции, являющийся базовым для всех потоков вещества и энергии в водной экосистеме. Оптическая активность пигментов, содержащихся в клетках фитопланктона, во многом формирует свойства гидрооптических полей. Это явление служит основой разработки оптических методов исследования распределения и свойств полей фитопланктона (Семовский, 1999). В частности, оптические свойства пигментов определяют возможности дистанционного исследования водных экосистем с помощью пассивных многоспектральных оптических сканеров, предназначенных для использования на самолетах и спутниках. В последнее время совокупность методов исследования водных экосистем с помощью оптических методов и связанный с этим анализ влияния биологических объектов на формирование подводных полей освещенности выделились в особую область океанологии и лимнологии, использующую методы гидрооптики и гидрофизики, с одной стороны, и гидробиологии, биофизики, теории популяций, с другой.
Модели, оперирующие с такими обобщенными таксономическими классами, как фитопланктон, допускают проверку с использованием систематических наблюдений, проводимых достаточно точными оптическими и флуоресцентными методами. Модельный подход по существу представляет собой еще одно средство анализа данных наблюдений.
Используя достаточно простые общие схемы трофических взаимодействий и адекватную модель первичной продукции, удается достаточно хорошо описать сезонную и географическую изменчивость полей пигментов фитопланктона, наблюдаемую в природе. Для построения подходящей модели первичной продукции, учитывающей специфику водоема, следует построить описание подводных полей освещенности в спектральном диапазоне фотосинтетически активной радиации (ФАР), что особенно важно при исследовании прибрежных районов и озер. В качестве первого приближения для этого можно использовать статистическую модель формирования поля подводной освещенности, то есть модель формирования спектрального поля ослабления света при известной вертикальной структуре распределения оптически активных компонентов. При наличии модели динамики биооптических полей возможна постановка задачи численного усваивания данных массовых наблюдений в моделях динамики экосистем. Речь в данном случае может идти как об усваивании измерений оптических характеристик, так и об усваивании таких оцененных по оптическим измерениям параметров как концентрация хлорофилла, взвешенных веществ и т. п. В практику физической океанографии и метеорологии в течение последних лет вошли методы численного усваивания данных наблюдений, основанные на идеях Г.И. Марчука. Такой подход является связующим звеном между анализом данных наблюдений и моделями природных явлений, для которых результаты расчетов далеко не всегда точно соответствуют динамике процессов, происходящих в природе. Методы численного усваивания (ассимиляции) наблюдений действуют как динамическая интерполяция и динамическое сглаживание данных, с одной стороны, а с другой, - позволяют адаптировать неизвестные или оцененные по ограниченным выборкам параметры моделей для получения в результате лучшего соответствия с действительностью. Поэтому представляется логичным поставить задачу о численном усваивании данных наблюдений в моделях динамики экосистем.
Имеются два различных подхода к оценке ФПП в водных экосистемах. Один из них получил широкое распространение, в частности, в трудах школы Г.Г. Винберга и основан на зависимости ассимиляционного числа ![]() от падающей ФАР. При таком подходе концентрация хлорофилла и падающая ФАР являются необходимыми исходными величинами. Другой подход к оценке продукции основан на зависимости эффективности фотосинтеза (квантового выхода)
от падающей ФАР. При таком подходе концентрация хлорофилла и падающая ФАР являются необходимыми исходными величинами. Другой подход к оценке продукции основан на зависимости эффективности фотосинтеза (квантового выхода) ![]() от падающей радиации
от падающей радиации ![]() . Для вычисления величины ФПП требуется информация о дневной дозе фотосинтетически усвоенной радиации
. Для вычисления величины ФПП требуется информация о дневной дозе фотосинтетически усвоенной радиации ![]() . Для этого требуется оценка спектра специфического поглощения света фитопланктоном
. Для этого требуется оценка спектра специфического поглощения света фитопланктоном ![]() и спектра падающей ФАР
и спектра падающей ФАР ![]() . Второй метод является несколько более сложным, но вместе с тем и более предпочтительным для использования, так как поглощение света фитопланктоном является точной и непосредственной характеристикой фотосинтетического аппарата, подверженной меньшей изменчивости, а его корреляция с первичной продукцией выше, чем для ассимиляционного числа. Однако применение этого метода пока ограничивается скудностью оценок спектра поглощения фитопланктона.
. Второй метод является несколько более сложным, но вместе с тем и более предпочтительным для использования, так как поглощение света фитопланктоном является точной и непосредственной характеристикой фотосинтетического аппарата, подверженной меньшей изменчивости, а его корреляция с первичной продукцией выше, чем для ассимиляционного числа. Однако применение этого метода пока ограничивается скудностью оценок спектра поглощения фитопланктона.
Математическое описание моделей.
1. Простые модели динамики фитопланктона.
Динамика фитопланктона, зоопланктона и растворенных биогенных веществ в верхнем слое океана.
![]() , (1.90.1)
, (1.90.1)
![]()
![]() , (1.90.2)
, (1.90.2)
![]() . (1.90.3)
. (1.90.3)
Здесь использованы следующие переменные: ![]() - фотосинтетическая продукция (описание ее расчета будет дано ниже);
- фотосинтетическая продукция (описание ее расчета будет дано ниже); ![]() - выедание (потребление) фитопланктона зоопланктоном;
- выедание (потребление) фитопланктона зоопланктоном; ![]() - концентрации фитопланктона, зоопланктона и биогенов в верхнем перемешанном слое (ВПС) (все в единицах азота [мгN/м-3]);
- концентрации фитопланктона, зоопланктона и биогенов в верхнем перемешанном слое (ВПС) (все в единицах азота [мгN/м-3]); ![]() - концентрации фитопланктона, зоопланктона и биогенов на нижней границе ВПС (в единицах азота [мгN/м-3]);
- концентрации фитопланктона, зоопланктона и биогенов на нижней границе ВПС (в единицах азота [мгN/м-3]); ![]() ,
, ![]() - интенсивность смертности фитопланктона и зоопланктона соответственно [с-1];
- интенсивность смертности фитопланктона и зоопланктона соответственно [с-1]; ![]() -неассимилированная фракция рациона хищника [безразм.]; h - глубина перемешанного слоя. Выедание описывается по формуле Ивлева
-неассимилированная фракция рациона хищника [безразм.]; h - глубина перемешанного слоя. Выедание описывается по формуле Ивлева
![]() , (1.91)
, (1.91)
где ![]() - максимальная интенсивность выедания хищником [с-1];
- максимальная интенсивность выедания хищником [с-1]; ![]() - константа Ивлева для выедания [м3 мгN-1]. Типичные значения основных параметров могут быть заимствованы, например, из работы (Wroblewski, 1989):
- константа Ивлева для выедания [м3 мгN-1]. Типичные значения основных параметров могут быть заимствованы, например, из работы (Wroblewski, 1989): ![]() = 0.3,
= 0.3, ![]() = 0.5,
= 0.5, ![]() = 0.1 сутки-1 = 0.1?1.157 10-5 c-1,
= 0.1 сутки-1 = 0.1?1.157 10-5 c-1, ![]() = 0.04?1.157 10-5 c-1.
= 0.04?1.157 10-5 c-1.
Для исследования процессов синоптического масштаба времени и пространства (дни, десятки километров) мы не имеем права использовать эти соотношения без учета стохастического фактора, описывающего изменчивость процессов вертикального перемешивания. В многочисленных работах было показано, что глубина перемешанного слоя и интенсивность вертикального перемешивания ![]() являются коротко коррелированными случайными процессами со значительной степенью изменчивости. Представим эти две переменные в виде сумм медленно- и быстроменяющейся компонент:
являются коротко коррелированными случайными процессами со значительной степенью изменчивости. Представим эти две переменные в виде сумм медленно- и быстроменяющейся компонент:
![]() , (1.92)
, (1.92)
где ![]() - среднемесячная климатическая глубина ВПС,
- среднемесячная климатическая глубина ВПС, ![]() - среднемесячная интенсивность вертикального перемешивания, вычисленная из климатических данных,
- среднемесячная интенсивность вертикального перемешивания, вычисленная из климатических данных, ![]() - случайный
- случайный ![]() - коррелированный процесс (белый шум) с дисперсией
- коррелированный процесс (белый шум) с дисперсией ![]() ,
, ![]() . С учетом этого система уравнений (1.90.1) - (1.90.3) примет следующий вид:
. С учетом этого система уравнений (1.90.1) - (1.90.3) примет следующий вид:
![]() , (1.93.1)
, (1.93.1)
![]()
![]() , (1.93.2)
, (1.93.2)
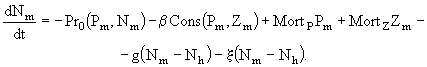 (1.93.3)
(1.93.3)
Здесь ![]() ,
, ![]() - случайный
- случайный ![]() - коррелированный процесс с дисперсией
- коррелированный процесс с дисперсией ![]() .
.
2. Модификация для эвтрофицированных водоемов.
Для водоемов, находящихся под значительным антропогенным воздействием, равно как и для находящихся в состоянии естественной эвтрофикации (благодаря выносу биогенных элементов реками), влияние растворенных ОВ в воде на формирование гидрооптических полей может быть весьма значительным. РОВ может поступать в систему как аллохтонное РОВ (вынос рек и перемешивание осадков) и как автохтонное РОВ (распад биологических остатков и детрита). Основными потребителями РОВ в водной экосистеме являются бактерии. В первом приближении динамика РОВ в экосистеме может быть описана соотношением
![]() , (2.94)
, (2.94)
где ![]() - средняя концентрация РОВ в ВПС (в единицах азота [мгN/м-3]);
- средняя концентрация РОВ в ВПС (в единицах азота [мгN/м-3]); ![]() - интенсивность образования РОВ [с-1];
- интенсивность образования РОВ [с-1]; ![]() - концентрация РОВ на нижней границе ВПС (в единицах азота [мгN/м-3]);
- концентрация РОВ на нижней границе ВПС (в единицах азота [мгN/м-3]); ![]() - интенсивность потребления РОВ бактериями [с-1]. Оценки параметров (получены путем анализа экспериментального материала):
- интенсивность потребления РОВ бактериями [с-1]. Оценки параметров (получены путем анализа экспериментального материала): ![]() = 0.71?1.157 10-6 c-1,
= 0.71?1.157 10-6 c-1, ![]() - 0.409?1.157 10-10 c-1.
- 0.409?1.157 10-10 c-1.
3. Трехмерная модель без вертикальной структуры.
![]() , (3.95.1)
, (3.95.1)
![]()
![]() , (3.95.2)
, (3.95.2)
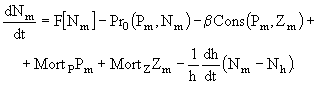 (3.95.3)
(3.95.3)
Оператор F[?] может быть взят в одной из следующих форм:
![]() - изотропная горизонтальная диффузия в отсутствие адвекции,
- изотропная горизонтальная диффузия в отсутствие адвекции, ![]() - коэффициент турбулентной диффузии;
- коэффициент турбулентной диффузии; ![]() - двумерная горизонтальная диффузия в отсутствие адвекции,
- двумерная горизонтальная диффузия в отсутствие адвекции, ![]() - тензор турбулентной диффузии;
- тензор турбулентной диффузии; ![]() - двумерная горизонтальная диффузия и адвекция в поле скоростей
- двумерная горизонтальная диффузия и адвекция в поле скоростей ![]() , полученном из наблюдений или из гидродинамической модели.
, полученном из наблюдений или из гидродинамической модели.
4. Моделирование вертикальной динамики экосистемы.
Данной системой уравнений описывается крупномасштабная изменчивость поля фитопланктона.
![]() , (4.96.1)
, (4.96.1)
![]()
![]() , (4.96.2)
, (4.96.2)
![]() (4.96.3)
(4.96.3)
![]() - кинематический коэффициент вертикальной турбулентной диффузии;
- кинематический коэффициент вертикальной турбулентной диффузии; ![]() - скорость гравитационного оседания клеток фитопланктона и зоопланктонных организмов.
- скорость гравитационного оседания клеток фитопланктона и зоопланктонных организмов.
5. Изменчивость соотношений между содержанием углерода, хлорофилла и азота в клетках фитопланктона.
При моделировании в данной работе используются переменные значения параметров Chl_a, C. Зависимость между этими величинами задается формулой
![]() .
.
Эта формула получена на основании данных непосредственных наблюдений. Атомное соотношение C/N взято постоянным и равным 5.
6. Простейшая модель усредненной продукции в столбе воды.
В данной работе не рассматриваются соотношения, используемые в основной массе моделей морских и озерных систем и описывающие ФПП как прямо пропорциональную концентрации фитопланктона в деятельном слое. При таком подходе не могут быть учтены вопросы эффектов самозатенения фитопланктона, лимитирования светом, влияния других оптически важных факторов, специфики географических условий. Одна из простых моделей ФПП, использованная в данной работе, предложена в выше упомянутой работе (Wroblewski, 1989):
![]() , (6.97)
, (6.97)
где ![]() - ФПП в единицах ассимилированного азота [мгN м-3 с-1];
- ФПП в единицах ассимилированного азота [мгN м-3 с-1]; ![]() - максимальная интенсивность роста фитопланктона [с-1];
- максимальная интенсивность роста фитопланктона [с-1]; ![]() - постоянная полунасыщения [мгN м-3], равная 0.2 (Wroblewski, 1989).
- постоянная полунасыщения [мгN м-3], равная 0.2 (Wroblewski, 1989).
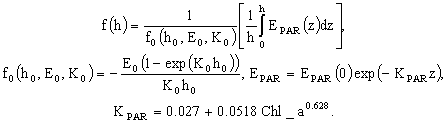 (6.98)
(6.98)
Здесь ![]() - климатическая глубина перемешанного слоя [м];
- климатическая глубина перемешанного слоя [м]; ![]() - осредненная за день величина
- осредненная за день величина ![]() - рассеянной фотосинтетически активной радиации (ФАР) на глубине
- рассеянной фотосинтетически активной радиации (ФАР) на глубине ![]() [Ватт м-2];
[Ватт м-2]; ![]() - климатический коэффициент вертикального ослабления для ФАР;
- климатический коэффициент вертикального ослабления для ФАР; ![]() - осредненная за день ФАР на глубине z [Ватт м-2];
- осредненная за день ФАР на глубине z [Ватт м-2]; ![]() - осредненный по глубине и за день коэффициент вертикального ослабления рассеянной падающей ФАР.
- осредненный по глубине и за день коэффициент вертикального ослабления рассеянной падающей ФАР. ![]() и
и ![]() измеряются в [м-1]. В работе (Wroblewski, 1989) для Северной Атлантики приведены следующие средние значения коэффициентов:
измеряются в [м-1]. В работе (Wroblewski, 1989) для Северной Атлантики приведены следующие средние значения коэффициентов: ![]() = 10,
= 10, ![]() = 2.1?1.157 10-6c-1,
= 2.1?1.157 10-6c-1, ![]() = 0,05;
= 0,05; ![]() = 125. Все они могут претерпевать значительную внутрисезонную изменчивость. Для окраинных морей, прибрежных вод и озер
= 125. Все они могут претерпевать значительную внутрисезонную изменчивость. Для окраинных морей, прибрежных вод и озер ![]() рассчитывают с учетом изменения оптических свойств воды за счет дополнительных поступлений РОВ и других активных компонентов, не связанных статистически с концентрацией хлорофилла:
рассчитывают с учетом изменения оптических свойств воды за счет дополнительных поступлений РОВ и других активных компонентов, не связанных статистически с концентрацией хлорофилла:
![]() (6.99)
(6.99)
где величина ![]() зависит от региона и сезона.
зависит от региона и сезона.
7. Спектральная модель оптических полей и первичной продукции.
Входными параметрами для модели являются вертикальный профиль хлорофилла ![]() [мг м-3], спектр падающей рассеянной освещенности, поглощенной в воде [мЕ м-2], и температура воды Т [°С]. Для представления поля подводной освещенности используется представление спектра коэффициента ослабления рассеянной падающей освещенности
[мг м-3], спектр падающей рассеянной освещенности, поглощенной в воде [мЕ м-2], и температура воды Т [°С]. Для представления поля подводной освещенности используется представление спектра коэффициента ослабления рассеянной падающей освещенности ![]() и его составляющей
и его составляющей ![]() , связанной с ослаблением света клетками фитопланктона [м-1] в зависимости от концентрации хлорофилла Chl_a в следующем виде:
, связанной с ослаблением света клетками фитопланктона [м-1] в зависимости от концентрации хлорофилла Chl_a в следующем виде:
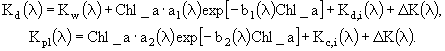 (7.100)
(7.100)
Параметры ![]() , а также коэффициент ослабления чистой воды находят по результатам измерений (В частности, для океанских и ряда морских вод существуют таблицы этих величин).
, а также коэффициент ослабления чистой воды находят по результатам измерений (В частности, для океанских и ряда морских вод существуют таблицы этих величин).
Спектр падающей рассеянной освещенности радиации в диапазоне ФАР на глубине z можно вычислить, используя известный спектр ослабления на каждой глубине и величину освещенности на поверхности по формуле
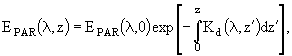 (7.101)
(7.101)
а энергию, поглощенную фитопланктоном, по формуле
 (7.102)
(7.102)
Зависимость квантового выхода фотосинтеза Ф [квант?атомС-1] от падающей подводной освещенности в диапазоне ФАР ![]() , концентрации хлорофилла на поверхности и температуры эвфотического слоя описывается соотношениями (приведены в общем виде):
, концентрации хлорофилла на поверхности и температуры эвфотического слоя описывается соотношениями (приведены в общем виде):
 (7.103)
(7.103)
где m, n, p, q - параметры, ![]() = 6.4?1019 [квант м-1 с-1],
= 6.4?1019 [квант м-1 с-1], ![]() = 1.77.
= 1.77.
На основе концепции экосистемного динамического моделирования (системной экодинамики) проведено изучение изменчивости оптических компонентов и полей концентраций растворенных и взвешенных веществ для прибрежной части Балтийского моря и оз. Байкал (Semovski, 1999). В качестве биологических переменных в модели рассматривались фитопланктон, зоопланктон и бактериопланктон. При моделировании полей подводной освещенности учитывались показатели ослабления чистой воды, поглощение света пигментами хлорофилла, оптические свойства "желтого вещества", ассоциированного с растворенным органическим веществом и две фракции взвешенного вещества. Моделирование выполнялось для изучения весенне-летнего цветения фитопланктона в Гданьском заливе (южная часть Балтийского моря) вдоль разреза и для описания годовых изменений в водной экосистеме оз. Байкал.
Дополнительная информация:
© 2001-2026 Кафедра биофизики МГУ