Для того, чтобы количественно оценить роль зоопланктона в круговороте органического вещества водоемов, необходимо знать не только численность и биомассу отдельных популяций, но и скорость продуцирования ими органического вещества, степень использования этой продукции хищными видами зоопланктона и рыбами, вклад каждой популяции в общую продукцию зоопланктона. При этом чрезвычайно важно учитывать изменения этих продукционных характеристик в течение года в зависимости от конкретных условий. Провести всестороннее исследование зоопланктона, основываясь только на результатах обработки стандартных проб, которые к тому же и берутся нерегулярно, невозможно. В этой ситуации методы математического моделирования оказываются единственной возможностью анализа и попыткой поиска ответов на многие конкретные вопросы гидробиологии.
Основные принципы, на основе которых строятся модели зоопланктонных сообществ, состоят в следующем. В основе моделей лежит общее уравнение, определяющее изменение за единицу времени численности популяции N как разность между количествами Q и M особей, соответственно поступивших в популяцию и выбывших из нее по разным причинам (Приходько, 1976):
![]() . (1)
. (1)
Величины Q и M вычисляются отдельно с учетом влияющих на них факторов, как внутрипопуляционных, так и внешних. Одной из главных характеристик популяции в каждый момент времени является ее возрастной состав, не зная которого, нельзя правильно оценить биомассу популяции, число родившихся и выбывших особей, продукцию популяции. Чтобы учесть в модели изменение возрастного состава, особи популяции разбиваются на несколько групп по какому-либо признаку, присущему всем особям, например, по возрасту, весу, размеру или стадии развития, спектру питания, воздействию хищников. Соответственно уравнение (1) применяется не ко всей популяции в целом, а к каждой ее группе в отдельности. Получается система уравнений
![]() (2)
(2)
или
![]() , (3)
, (3)
где k - число групп, ![]() - численность i-ой группы в момент времени t. Число поступивших особей
- численность i-ой группы в момент времени t. Число поступивших особей ![]() определяется для первой группы (самой младшей) скоростью размножения, а для остальных - скоростью развития особей, то есть длительностью их пребывания в предыдущей группе. Число выбывших из группы
определяется для первой группы (самой младшей) скоростью размножения, а для остальных - скоростью развития особей, то есть длительностью их пребывания в предыдущей группе. Число выбывших из группы ![]() определяется скоростью развития особей и их смертностью.
определяется скоростью развития особей и их смертностью.
Длительность ![]() развития особи в каждой группе зависит от ее биологических особенностей, температуры среды
развития особи в каждой группе зависит от ее биологических особенностей, температуры среды ![]() и условий питания, которые можно охарактеризовать степенью удовлетворения пищевых потребностей особи
и условий питания, которые можно охарактеризовать степенью удовлетворения пищевых потребностей особи ![]() . Таким образом, если
. Таким образом, если ![]() - продолжительность развития при оптимальных условиях, то есть при
- продолжительность развития при оптимальных условиях, то есть при ![]() и температуре
и температуре ![]() , то
, то
![]() . (4)
. (4)
Конкретный вид функции зависит от особенностей развития особей изучаемой популяции.
Пищевые потребности (максимальный рацион) ![]() особи весом
особи весом ![]() вычисляются из предположения о том, что ассимилированной пищи должно хватить на возмещение энергетических затрат организма в процессе обмена
вычисляются из предположения о том, что ассимилированной пищи должно хватить на возмещение энергетических затрат организма в процессе обмена ![]() и на максимальный весовой прирост
и на максимальный весовой прирост ![]() :
:
![]() , (5)
, (5)
где ![]() ,
, ![]() - усвояемость пищи. Величины
- усвояемость пищи. Величины ![]() можно рассчитать по экспериментально установленной зависимости
можно рассчитать по экспериментально установленной зависимости ![]() , в которую следует вводить поправку на температуру. Просуммировав максимальные рационы всех особей популяции, получим пищевые потребности (максимальный рацион) всей популяции
, в которую следует вводить поправку на температуру. Просуммировав максимальные рационы всех особей популяции, получим пищевые потребности (максимальный рацион) всей популяции ![]() , сопоставлением которых с доступным количеством пищи S и определяются величины
, сопоставлением которых с доступным количеством пищи S и определяются величины ![]() . В частности, можно воспользоваться модифицированной формулой В. С. Ивлева (Ивлев, 1955)
. В частности, можно воспользоваться модифицированной формулой В. С. Ивлева (Ивлев, 1955)
![]() , (6)
, (6)
где ![]() - эмпирический коэффициент. Очевидно, что если все особи популяции питаются одним и тем же кормом,
- эмпирический коэффициент. Очевидно, что если все особи популяции питаются одним и тем же кормом, ![]() будет одинаковыми для всех групп.
будет одинаковыми для всех групп.
Зная реальную продолжительность ![]() развития особи в каждой группе, можно определить число особей
развития особи в каждой группе, можно определить число особей ![]() , переходящих в момент времени t в следующую группу. Для
, переходящих в момент времени t в следующую группу. Для ![]() особей, пришедших в i-ую группу в момент времени
особей, пришедших в i-ую группу в момент времени ![]() , рассчитывается величина
, рассчитывается величина
![]() , (7)
, (7)
которую можно интерпретировать как степень развития в i-ой группе, достигнутую особями к моменту времени t. Считается, что переход этих особей в группу ![]() осуществляется в тот момент Т, когда
осуществляется в тот момент Т, когда ![]() . Таким образом,
. Таким образом,
![]() , (8)
, (8)
где ![]() - смертность особей i-ой группы.
- смертность особей i-ой группы.
Особи популяции гибнут по нескольким причинам. Они подвергаются воздействию хищников, могут умирать от недостатка пищи и не исключена возможность естественной их гибели от старости, паразитов или болезней. Если ![]() - часть рациона хищников, приходящаяся на особей i-ой группы, а
- часть рациона хищников, приходящаяся на особей i-ой группы, а ![]() - биомасса этой группы, то коэффициент смертности от воздействия хищников при отсутствии других причин гибели можно представить, например, в виде
- биомасса этой группы, то коэффициент смертности от воздействия хищников при отсутствии других причин гибели можно представить, например, в виде
![]() . (9)
. (9)
Формула (9) учитывает зависимость степени выедания зоопланктона как от количества самого зоопланктона, так и от рациона хищников и отражает тот факт, что с увеличением плотности жертв или рациона хищников смертность возрастает и стремится к какому-то предельному значению ![]() , благодаря чему полное выедание жертвы невозможно (Ивлев, 1955). Гибель от голода может наступать только в том случае, если особь потребляет энергии меньше, чем тратит. Определив критическую степень удовлетворения пищевых потребностей
, благодаря чему полное выедание жертвы невозможно (Ивлев, 1955). Гибель от голода может наступать только в том случае, если особь потребляет энергии меньше, чем тратит. Определив критическую степень удовлетворения пищевых потребностей ![]() , при которой вся полученная с пищей энергия расходуется на обмен, можно считать, что коэффициент смертности от голода равен
, при которой вся полученная с пищей энергия расходуется на обмен, можно считать, что коэффициент смертности от голода равен
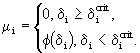 (10)
(10)
Например, можно принять, что ![]() . Так как обе эти причины могут действовать одновременно, то коэффициент общей смертности определяется по формуле
. Так как обе эти причины могут действовать одновременно, то коэффициент общей смертности определяется по формуле
![]() . (11)
. (11)
Естественную смертность можно рассматривать как выход старших особей из этой группы после завершения всего цикла развития.
Размножение особей и тем самым пополнение первой группы вычисляется по количеству яиц ![]() , отложенных в популяции за единицу времени:
, отложенных в популяции за единицу времени:
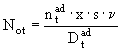 , (12)
, (12)
где ![]() ,
, ![]() - соответственно число и продолжительность развития половозрелых особей, x - соотношение полов, s - число кладок у самки в течение ее жизни,
- соответственно число и продолжительность развития половозрелых особей, x - соотношение полов, s - число кладок у самки в течение ее жизни, ![]() - количество яиц в кладке в расчете на половозрелую самку. Смертность яиц, вынашиваемых самками, принимается равной смертности взрослых самок.
- количество яиц в кладке в расчете на половозрелую самку. Смертность яиц, вынашиваемых самками, принимается равной смертности взрослых самок.
Таким образом, величины ![]() и
и ![]() в формуле (3) равны
в формуле (3) равны
![]() . (13)
. (13)
Если в каждый момент времени известны численность и средний вес ![]() особей каждой группы, то биомасса в момент времени
особей каждой группы, то биомасса в момент времени ![]() и продукция популяции за время
и продукция популяции за время ![]() вычисляется по формулам
вычисляется по формулам
![]() , (14)
, (14)
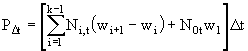 . (15)
. (15)
Для того чтобы применить такой подход к изучению какой-либо конкретной популяции зоопланктона, надо знать следующие величины: температуру воды места обитания ![]() , биомассу кормовых объектов S, рацион хищников V и его распределение по выделенным группам рассматриваемой популяции, продолжительность развития особей в каждой группе при оптимальных условиях
, биомассу кормовых объектов S, рацион хищников V и его распределение по выделенным группам рассматриваемой популяции, продолжительность развития особей в каждой группе при оптимальных условиях ![]() , средний вес особи каждой группы
, средний вес особи каждой группы ![]() , плодовитость
, плодовитость ![]() , соотношение полов x, а также численности групп в начальный момент времени. При этом модель не ограничивает возможности дальнейшей конкретизации структуры популяции и условий ее обитания.
, соотношение полов x, а также численности групп в начальный момент времени. При этом модель не ограничивает возможности дальнейшей конкретизации структуры популяции и условий ее обитания.
Описанная модель была использована для изучения некоторых закономерностей продукционного процесса популяций трех доминирующих видов зоопланктона оз. Дальнего (Камчатка) - Neutrodiaptomus angustilobus Sars, Cyclops scutifer Sars и Daphnia longiremis Sars (Приходько, 1976; Приходько, 1978). В частности, было показано, что при изменении рационов потребителей зоопланктона все показатели продукционного процесса рассмотренных популяций изменяются в более узких пределах, чем при изменении количества доступного корма, то есть для зоопланктонной популяции условия питания имеют большее значение, чем воздействие хищников. Такой характер влияния кормовых условий и пресса хищников на популяции планктонных ракообразных подтверждается как многолетними наблюдениями на оз. Дальнем, так и многочисленными исследованиями на других водоемах.
Три популяции планктонных ракообразных оз. Дальнего - Neutrodiaptomus angustilobus Sars, Cyclops scutifer Sars и Daphnia longiremis Sars - составляют основную долю биомассы зоопланктона озера и тесно связаны между собой. У них общие источники питания и хищник. Кроме того, старшие копеподиты циклопов являются хищниками по отношению к науплиусам и младшим копеподитам своей популяции и популяции диаптомусов. Следовательно, как потребление пищи, так и гибель особей каждой популяции определяются численностью и возрастным составом не только ее самой, но и других популяций. Поэтому очевидно, что все три популяции должны рассматриваться как элементы модели зоопланктона озера. Источники питания основной массы зоопланктона - фитопланктон, бактерии, простейшие, детрит, конечно, испытывают воздействие со стороны рассматриваемых популяций. Чтобы излишне не усложнять модель, динамика кормовой базы задавалась как один из входов модели. То же можно сказать и о другом внешнем для зоопланктона факторе - рыбах - планктофагах.
Каждая популяция была разбита на следующие группы (Приходько, 1978). Neutrodiaptomus angustilobus Sars: ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() ; Cyclops scutifer Sars:
; Cyclops scutifer Sars: ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() ; Daphnia longiremis Sars:
; Daphnia longiremis Sars: ![]() ,
, ![]() ,
, ![]() ,
, ![]() . По источникам питания все ракообразные делятся на мирных - питающихся фитопланктоном и бактериями (S), и хищных, в спектр питания которых добавляется мирный зоопланктон. К мирным относятся вторая - пятая группы популяций диаптомусов (
. По источникам питания все ракообразные делятся на мирных - питающихся фитопланктоном и бактериями (S), и хищных, в спектр питания которых добавляется мирный зоопланктон. К мирным относятся вторая - пятая группы популяций диаптомусов (![]() ,
, ![]() ,
, ![]() ,
, ![]() ), вторая - третья группы популяций циклопов (
), вторая - третья группы популяций циклопов (![]() ,
, ![]() ) и третья - четвертая группы популяции дафний (
) и третья - четвертая группы популяции дафний (![]() ,
, ![]() ). Группу хищных составляют копеподиты IV - V стадий и взрослые особи популяции циклопов (
). Группу хищных составляют копеподиты IV - V стадий и взрослые особи популяции циклопов (![]() ,
, ![]() ), которые часть своего рациона
), которые часть своего рациона ![]() удовлетворяют за счет пищи S, другую
удовлетворяют за счет пищи S, другую ![]() - за счет второй и третьей групп популяций диаптомусов и циклопов (
- за счет второй и третьей групп популяций диаптомусов и циклопов (![]() ,
, ![]() ,
, ![]() ,
, ![]() ). Пищевые потребности рыб (V) распределяются между всеми популяциями.
). Пищевые потребности рыб (V) распределяются между всеми популяциями.
Как и модели изолированных популяций, модель зоопланктонного сообщества представляет собой систему конечно-разностных уравнений, описывающих изменения численности каждой группы рассматриваемых популяций за единицу времени. Пополнение группы осуществляется в результате размножения (для группы яиц) и перехода особей из одной группы в другую в процессе роста. Число яиц, откладываемых в каждой популяции в данный момент, определяли с учетом изменяющихся во времени средней плодовитости самок, длительности периодов между кладками, соотношения полов и числа способных к размножению особей каждой популяции. Скорость роста особей, то есть их перехода в следующую группу, выражается в модели через реальную продолжительность их развития в каждой группе. Длительность ![]() развития особи зависит от ее биологических особенностей, температуры среды
развития особи зависит от ее биологических особенностей, температуры среды ![]() и условий питания, которые можно охарактеризовать степенью удовлетворения пищевых потребностей особи
и условий питания, которые можно охарактеризовать степенью удовлетворения пищевых потребностей особи ![]() . Таким образом, если
. Таким образом, если ![]() - продолжительность развития при оптимальных условиях, то есть при
- продолжительность развития при оптимальных условиях, то есть при ![]() и температуре
и температуре ![]() , то
, то ![]() . Конкретный вид функции зависит от особенностей развития особей изучаемой популяции.
. Конкретный вид функции зависит от особенностей развития особей изучаемой популяции. ![]() определяется путем сопоставления суммарной потребности в пище всех особей каждого вида с имеющимся количеством этой пищи. Пищевые потребности (максимальный рацион)
определяется путем сопоставления суммарной потребности в пище всех особей каждого вида с имеющимся количеством этой пищи. Пищевые потребности (максимальный рацион) ![]() особи i-ой группы со средним весом
особи i-ой группы со средним весом ![]() вычисляются, как указывалось выше, из предположения о том, что ассимилированной пищи должно хватить на возмещение энергетических затрат организма в процессе обмена
вычисляются, как указывалось выше, из предположения о том, что ассимилированной пищи должно хватить на возмещение энергетических затрат организма в процессе обмена ![]() и на максимальный весовой прирост
и на максимальный весовой прирост ![]() :
:
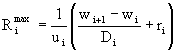 , (16)
, (16)
где ![]() ,
, ![]() - коэффициент усвоения пищи. Величины
- коэффициент усвоения пищи. Величины ![]() можно рассчитать по экспериментально установленной зависимости
можно рассчитать по экспериментально установленной зависимости ![]() , в которую следует вводить поправку на температуру. Суммарная потребность зоопланктона в пище, состоящей из фитопланктона и бактерий, равна
, в которую следует вводить поправку на температуру. Суммарная потребность зоопланктона в пище, состоящей из фитопланктона и бактерий, равна
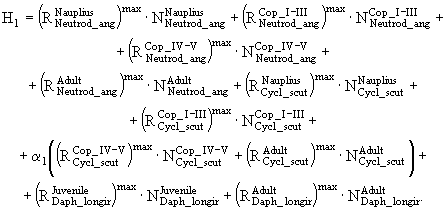 (17)
(17)
Потребность хищных циклопов в животной пище определяется соотношением
![]() . (18)
. (18)
В модели предполагается, что рыбы потребляют только особей старшего возраста каждой популяции и не являются поэтому пищевыми конкурентами хищных циклопов.
Модифицируя формулу В. С. Ивлева (Ивлев, 1955), получим степень удовлетворения потребности в пище S всех питающихся ею особей (![]() ):
):
![]() , (19)
, (19)
где ![]() . Аналогично, если Q - суммарная биомасса науплиусов и младших копеподитов популяций диаптомусов и циклопов, то
. Аналогично, если Q - суммарная биомасса науплиусов и младших копеподитов популяций диаптомусов и циклопов, то
![]() ,
, ![]() . ( 20)
. ( 20)
Таким образом, степень удовлетворения пищевых потребностей для мирных особей равна ![]() , для хищных особей -
, для хищных особей - ![]() .
.
Выбывание особей из каждой группы обусловлено переходом их в следующую группу (или гибелью от старости для взрослых особей каждой популяции) и гибелью под воздействием хищника или от голода. Предполагалось, что гибель особи от голода может наступить тогда, когда степень удовлетворения ее пищевых потребностей меньше критического значения ![]() , при котором вся полученная с пищей энергия расходуется на обмен. В данной модели было принято, что коэффициент смертности от голода равен:
, при котором вся полученная с пищей энергия расходуется на обмен. В данной модели было принято, что коэффициент смертности от голода равен:
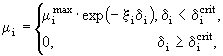 (21)
(21)
где ![]() - максимально возможная смертность,
- максимально возможная смертность, ![]() - эмпирический коэффициент. При моделировании гибели особей в результате воздействия хищников рацион рыб - V распределялся между популяциями диаптомусов, циклопов и дафний в долях
- эмпирический коэффициент. При моделировании гибели особей в результате воздействия хищников рацион рыб - V распределялся между популяциями диаптомусов, циклопов и дафний в долях ![]() ,
, ![]() и
и ![]() , а между особями i-ой группы - в долях
, а между особями i-ой группы - в долях ![]() . При этом предполагалось, что
. При этом предполагалось, что
![]() ;
; ![]() ;
; ![]() . (22)
. (22)
Распределение животной части рациона хищных циклопов ![]() между жертвами, то есть особями групп,
между жертвами, то есть особями групп, ![]() ,
, ![]() ,
, ![]() ,
, ![]() , предполагалось пропорциональным биомассе
, предполагалось пропорциональным биомассе ![]() ,
, ![]() ,
, ![]() ,
, ![]() этих групп. Таким образом, потребности хищников в особях каждой группы равны
этих групп. Таким образом, потребности хищников в особях каждой группы равны
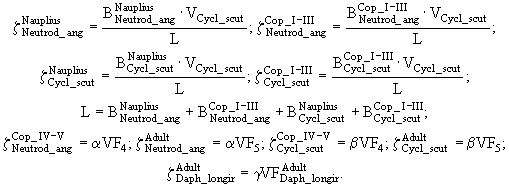 (23)
(23)
Смертность особей каждой группы от воздействия хищников, аналогично более ранним версиям модели, вычислялась по формуле
![]() , (24)
, (24)
где А - максимально возможная смертность, ![]() ,
, ![]() - эмпирические коэффициенты. При использовании этой формулы к биомассе взрослых особей добавляли биомассу яиц (исключая эфиппиумы в популяции дафний). Общую смертность особей i-ой группы считали равной
- эмпирические коэффициенты. При использовании этой формулы к биомассе взрослых особей добавляли биомассу яиц (исключая эфиппиумы в популяции дафний). Общую смертность особей i-ой группы считали равной
![]() . (25)
. (25)
Смертность яиц приравнивалась к смертности взрослых особей соответствующей популяции.
Данная модель зоопланктонного сообщества позволяет вычислять не только численность всех групп особей в каждый момент времени, но и такие характеристики, как биомасса, продукция и элиминация отдельной популяции, а также мирного и хищного зоопланктона озера в целом. При расчете этих величин биомассу и продукцию яиц циклопов включают в соответствующие характеристики хищного зоопланктона. Для популяции циклопов определяют как продукцию ее мирной и хищной части отдельно, так и продукцию всей популяции. Последнюю вычисляют с учетом факта потребления хищными циклопами собственной молоди.
В работах (Казанцева, Смирнова, 1985; Казанцева, Смирнова, 1986) рассматриваются математические модели зоопланктонного сообщества, которые построены на этих же принципах, но применительно к зоопланктонному сообществу южного, наиболее продуктивного, района Ладожского озера. Этот метод моделирования основан на подробном описании роста особей рассматриваемых популяций (с учетом возрастных групп) и их трофических взаимоотношений с учетом действия факторов внешней среды (температурные условия, обеспеченность пищей). Он позволяет получать как годовую динамику численности разных возрастных групп популяций, так и все необходимые продукционные характеристики каждой популяции в отдельности и всего сообщества в целом. В указанных работах для южного района Ладожского озера выделены четыре доминирующих популяции зоопланктона: Eudiaptomus gracilis Sars (мирный зоопланктон, питающийся фитопланктоном, бактериями и простейшими. Встречается круглый год, размножается с марта по сентябрь), Mesocyclops leuckarti Claus (факультативный хищник. Копеподиты IV-V стадий и половозрелые особи наряду с растительной пищей потребляют и животную - науплиусов и младших копеподитов Copepoda, мелких Cladocera, коловраток. В середине октября в развитии копеподитов IV-V стадий наступает диапауза, которая длится до момента освобождения озера ото льда (середина мая). В это время рост и питание особей прекращается. Популяция имеет два периода размножения - начало и конец лета), Bosmina obtusirostris lacustris Sars (мирный вид. На небольших глубинах наблюдается два максимума численности: в июне-июле и сентябре. Осенью в популяции появляются эфиппиальные самки, которые откладывают зимние яйца, развивающиеся весной) и Asplanchna priodonta Gosse (относится к хищникам. Взрослые особи едят мелких планктонных животных, в основном коловраток и простейших. С установлением ледяного покрова эти животные исчезают из планктона и появляются вновь в мае, когда начинают развиваться яйца, зимовавшие в илу. На мелководье наблюдается два максимума численности: в июне-июле и сентябре). Входами модели являются значения биомассы фитопланктона, бактерий и простейших, мелких коловраток, рациона рыб и температуры воды, которые задавались в конце каждого месяца. Рационы рыб приняты произвольно. При задании продолжительности развития особей разных возрастных групп и плодовитости в расчете на половозрелую самку пользовались литературными данными либо принимались их предполагаемые значения. Шаг модели равен 2 суткам. Выходами служат годовая динамика численности каждой группы зоопланктона, суточные, месячные, сезонные и годовые значения биомассы, продукции, элиминации и коэффициентов P/B отдельных популяций, а также групп мирного и хищного зоопланктона. Основными критериями качества модели являлись устойчивость модельной системы, то есть повторяемость выходных величин из года в год при одних и тех же значениях входов, возврат численности групп в конце модельного года к начальным значениям и правдоподобность результатов.
Особи каждой популяции, включая яйца или зародыши, разбиты на возрастные группы: яйца, науплиусы, копеподиты I-III, копеподиты IV-V и взрослые (соответственно ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() для E. gracilis;
для E. gracilis; ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() для M. leuckarti); эфиппиумы, партеногенетические яйца, молодь и взрослые (соответственно
для M. leuckarti); эфиппиумы, партеногенетические яйца, молодь и взрослые (соответственно ![]() ,
, ![]() ,
, ![]() ,
, ![]() для B. obtusirostris); зимние и летние яйца, молодь и взрослые (соответственно
для B. obtusirostris); зимние и летние яйца, молодь и взрослые (соответственно ![]() ,
, ![]() ,
, ![]() ,
, ![]() для A. priodonta). Для всех групп заданы средние веса
для A. priodonta). Для всех групп заданы средние веса ![]() и длительность пребывания
и длительность пребывания ![]() особей в данной группе при 20°С при полном удовлетворении пищевых потребностей. По истечении срока пребывания особи в данной возрастной группе происходит ее переход в следующую группу. Скорость перехода зависит от температуры водной массы и условий питания, то есть
особей в данной группе при 20°С при полном удовлетворении пищевых потребностей. По истечении срока пребывания особи в данной возрастной группе происходит ее переход в следующую группу. Скорость перехода зависит от температуры водной массы и условий питания, то есть ![]() . Принималось, что продукцию особь образует именно в момент этого перехода. Кроме того, для популяции M. leuckarti предусмотрен уход старших копеподитов в диапаузу.
. Принималось, что продукцию особь образует именно в момент этого перехода. Кроме того, для популяции M. leuckarti предусмотрен уход старших копеподитов в диапаузу.
По источникам питания зоопланктон модельного сообщества был разделен на две группы: мирный зоопланктон (![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() ) и хищный зоопланктон (
) и хищный зоопланктон (![]() ,
, ![]() ,
, ![]() ). Объектами питания мирного зоопланктона являются фитопланктон (
). Объектами питания мирного зоопланктона являются фитопланктон (![]() ) и детрит совместно с бактериями (
) и детрит совместно с бактериями (![]() ). Хищный зоопланктон питается мелкими коловратками и простейшими (
). Хищный зоопланктон питается мелкими коловратками и простейшими (![]() ), а также группами
), а также группами ![]() ,
, ![]() ,
, ![]() ,
, ![]() (
(![]() ) и
) и ![]() ,
, ![]() ,
, ![]() (
(![]() ). Доля j-го вида пищи в рационе особи i-ой группы вычислялась как часть, которую данный вид пищи составляет в общей биомассе пищевых объектов особей этой группы. Пищевые потребности (максимальный рацион)
). Доля j-го вида пищи в рационе особи i-ой группы вычислялась как часть, которую данный вид пищи составляет в общей биомассе пищевых объектов особей этой группы. Пищевые потребности (максимальный рацион) ![]() особи i-ой группы со средним весом
особи i-ой группы со средним весом ![]() вычисляются из предположения о том, что ассимилированной пищи должно хватить на возмещение энергетических затрат организма в процессе обмена
вычисляются из предположения о том, что ассимилированной пищи должно хватить на возмещение энергетических затрат организма в процессе обмена ![]() и на максимальный весовой прирост
и на максимальный весовой прирост ![]() :
: 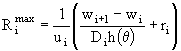 , где
, где ![]() ,
, ![]() - коэффициент усвоения пищи,
- коэффициент усвоения пищи, ![]() - температурная поправка. Степень удовлетворения потребностей в каждом виде пищи (
- температурная поправка. Степень удовлетворения потребностей в каждом виде пищи (![]() ) определяется сопоставлением суммарных потребностей в этой пище (
) определяется сопоставлением суммарных потребностей в этой пище (![]() ) и имеющегося ее количества (
) и имеющегося ее количества (![]() ). По величинам
). По величинам ![]() вычисляются реальные рационы (
вычисляются реальные рационы (![]() ) особей каждой группы и соответствующие степени удовлетворения пищевых потребностей
) особей каждой группы и соответствующие степени удовлетворения пищевых потребностей ![]() , которые определяют реальные продолжительности развития
, которые определяют реальные продолжительности развития ![]() при данных температурных и пищевых условиях.
при данных температурных и пищевых условиях.
В модели были предусмотрены три причины гибели зоопланктона: выедание (![]() ) его рыбами (V) и хищным зоопланктоном (
) его рыбами (V) и хищным зоопланктоном (![]() ,
, ![]() , а также
, а также ![]() ), гибель от недостатка пищи, когда потребленная с пищей энергия не восполняет трат на обмен (
), гибель от недостатка пищи, когда потребленная с пищей энергия не восполняет трат на обмен (![]() ), и естественная гибель после завершения всего цикла развития. Плодовитость самок каждой популяции по каждому виду яиц (в расчете на половозрелую самку), а также факторы внешней среды задавались как функции времени.
), и естественная гибель после завершения всего цикла развития. Плодовитость самок каждой популяции по каждому виду яиц (в расчете на половозрелую самку), а также факторы внешней среды задавались как функции времени.
Таким образом, модель сообщества популяции зоопланктона описывается следующими основными соотношениями. Как уже было сказано, траты на обмен зависят от веса особей i-ой группы и температуры воды ![]() :
:
![]() . (26)
. (26)
В частности, величины ![]() можно рассчитать по экспериментально установленной зависимости
можно рассчитать по экспериментально установленной зависимости
![]() . (27)
. (27)
Максимальный рацион рассчитывается с учетом коэффициента усвоения пищи ![]() :
:
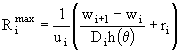 . (28)
. (28)
После этого могут быть определены суммарные потребности организмов i-ой группы в j-ом виде пищи:
![]() , (29)
, (29)
где ![]() соответствующие коэффициенты предпочтения организмами i-ой группы j-го вида пищи,
соответствующие коэффициенты предпочтения организмами i-ой группы j-го вида пищи, ![]() - численность i-ой группы,
- численность i-ой группы, ![]() - выедание рыбами пищи. Степень удовлетворения потребности в j-ом виде пищи всех питающихся ею особей (
- выедание рыбами пищи. Степень удовлетворения потребности в j-ом виде пищи всех питающихся ею особей (![]() ) задается соотношением, которое является модификацией формулы Ивлева:
) задается соотношением, которое является модификацией формулы Ивлева:
![]() , (30)
, (30)
где ![]() . Зная максимальные рационы и степень удовлетворения потребностей в определенном виде пищи, можно рассчитать реальные рационы:
. Зная максимальные рационы и степень удовлетворения потребностей в определенном виде пищи, можно рассчитать реальные рационы:
![]() (31)
(31)
и соответствующие степени удовлетворения пищевых потребностей
![]() . (32)
. (32)
Скорость перехода организмов в следующую возрастную группу, как уже было сказано, зависит от температуры водной массы и условий питания, то есть ![]() . В соответствии с этим
. В соответствии с этим
![]() . (33)
. (33)
Если ![]() - часть рациона хищников, приходящаяся на особей i-ой группы, а
- часть рациона хищников, приходящаяся на особей i-ой группы, а ![]() - биомасса этой группы, то коэффициент смертности от воздействия хищников при отсутствии других причин гибели можно представить, например, в виде
- биомасса этой группы, то коэффициент смертности от воздействия хищников при отсутствии других причин гибели можно представить, например, в виде
![]() . (34)
. (34)
Формула (34) учитывает зависимость степени выедания зоопланктона как от количества самого зоопланктона, так и от рациона хищников и отражает тот факт, что с увеличением плотности жертв или рациона хищников смертность возрастает и стремится к какому-то предельному значению ![]() , благодаря чему полное выедание жертвы невозможно (Ивлев, 1955). Гибель от голода может наступать только в том случае, если особь потребляет энергии меньше, чем тратит. Определив критическую степень удовлетворения пищевых потребностей
, благодаря чему полное выедание жертвы невозможно (Ивлев, 1955). Гибель от голода может наступать только в том случае, если особь потребляет энергии меньше, чем тратит. Определив критическую степень удовлетворения пищевых потребностей
![]() , (35)
, (35)
при которой вся полученная с пищей энергия расходуется на обмен, можно считать, что коэффициент смертности от голода равен
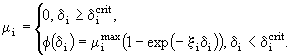 (36)
(36)
Так как обе эти причины могут действовать одновременно, то коэффициент общей смертности определяется по формуле
![]() . (37)
. (37)
Естественную смертность можно рассматривать как выход старших особей из этой группы после завершения всего цикла развития. Размножение особей и тем самым пополнение первой группы вычисляется по количеству яиц ![]() , отложенных в популяции за единицу времени:
, отложенных в популяции за единицу времени:
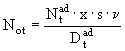 , (38)
, (38)
где ![]() ,
, ![]() - соответственно число и продолжительность развития половозрелых особей, x - соотношение полов, s - число кладок у самки в течение ее жизни,
- соответственно число и продолжительность развития половозрелых особей, x - соотношение полов, s - число кладок у самки в течение ее жизни, ![]() - количество яиц в кладке в расчете на половозрелую самку. Смертность яиц, вынашиваемых самками, принимается равной смертности взрослых самок. Биомасса той или иной популяции зоопланктона определяется суммой следующего вида:
- количество яиц в кладке в расчете на половозрелую самку. Смертность яиц, вынашиваемых самками, принимается равной смертности взрослых самок. Биомасса той или иной популяции зоопланктона определяется суммой следующего вида:
![]() , (39)
, (39)
а ее продукция - соотношением
![]() , (40)
, (40)
в котором ![]() - количество особей, перешедших из группы i в группу i + 1;
- количество особей, перешедших из группы i в группу i + 1; ![]() - генеративная продукция за время
- генеративная продукция за время ![]() , то есть
, то есть
![]() . (41)
. (41)
Зоопланктон Ладожского озера, характеризующегося большим разнообразием рельефных, гидрологических и биотических условий, не является однородным по всей акватории озера. Поэтому при моделировании зоопланктона целесообразно выделять такие районы озера, внутри которых сообщество популяций зоопланктона можно считать стабильным и характерным для данного района.
Центральный район Ладожского озера на большей части своей акватории имеет среднюю глубину 60 - 70 м, на севере района глубины превышают 100 м, на юге близки к 40 м. Объем водной массы района составляет около 50% объема всего озера. Этот район большую часть года заполнен озерной водой, прибрежные воды проникают сюда лишь после исчезновения весеннего термобара, то есть к концу июня - июлю. Термический режим района характеризуется более длинной по сравнению с прибрежными мелководными районами весной, коротким летом и продолжительным осенним периодом. Фитопланктон здесь как по видовому составу, так и количественно беднее, чем в прибрежных районах, хотя летом в отдельные периоды его продукция может превышать продукцию фитопланктона в Волховской губе.
По данным проб, которые отбирались на станциях продольного разреза центральной части озера в разные годы и разное время вегетационного сезона, зоопланктон этого района представлен различными видами Copepoda, Cladocera и Rotatoria. Наиболее массовыми видами являются два вида Calanoida - Eudiaptomus gracilis Sars и Limnocalanus macrurus Sars, два вида Cyclopoida - Mesocyclops oithonoides Sars и Cyclops abyssorum Sars и представитель ветвистоусых Bosmina obtusirostris lacustris Sars. Особи L. macrurus и C. abyssorum предпочитают низкие температуры с оптимумом 8 - 10°С, остальные виды из перечисленных являются сравнительно теплолюбивыми. Это обусловливает неоднородность распределения зоопланктона по глубине в течение сезона.
По ряду объективных причин получение надежной количественной оценки биомассы и продукции зоопланктона данного района для всего столба воды как в отдельные моменты времени, так и за вегетационный сезон или за год, является сложной проблемой. Для ее решения естественно использовать метод математического моделирования. Модель, которая описывает развитие каждой популяции с учетом ее биологических особенностей, положения в экосистеме, взаимодействия с другими элементами экосистемы, позволит получать обоснованные оценки различных характеристик как отдельных популяций, так и всего сообщества в целом в любой момент или за любой интервал времени.
Имитационная модель зоопланктонного сообщества центрального района Ладожского озера (Казанцева, Смирнова, 1996) построена по принципу, который хорошо зарекомендовал себя при моделировании зоопланктона (Приходько, 1976; Приходько, 1978; Кузеванова и др., 1978; Казанцева, Смирнова, 1985; Казанцева, Смирнова, 1986). В соответствии с ним каждая популяция разбивается на несколько групп по возрастному, трофическому или иному признаку. Внутри группы особи считаются одинаковыми по основным физиологическим свойствам и различаются только по времени их поступления в группу. На каждом шаге модели, соответствующем некоторому промежутку времени, вычисляется изменение численности каждой группы за это время. Это изменение складывается из поступления и убыли особей. Источником поступления служит откладка яиц и переход особей в процессе роста в данную возрастную группу из предыдущей. Убыль происходит в результате перехода особей в следующую группу или гибели по разным причинам. Скорость роста, то есть перехода из данной группы в следующую, определяется биологическими свойствами вида, а также зависит от температурных и трофических условий. Стимулом для сезонных вертикальных перемещений является выбор оптимальных условий обитания.
При моделировании развития популяций по такому принципу необходимо знать основные эколого-физиологические характеристики особей моделируемых популяций. К ним относятся изменение средней массы особи в процессе роста, скорость развития, то есть длительность каждой стадии развития при разных температурах, зависимость интенсивности обмена особи от массы тела и температуры, интенсивность размножения в разных условиях, спектр питания и его эффективность. Кроме того, надо знать характер взаимодействия между популяциями, образующими моделируемое сообщество, а также годовую динамику факторов внешней среды, влияющих на развитие зоопланктона.
Моделируемое сообщество зоопланктона представлено в данной модели в виде трехъярусной системы. Каждый ярус соответствует определенному слою, взятому в столбе воды под квадратным метром поверхности, причем отсчет глубин и ярусов ведется от дна к поверхности. В модели принимается, что III-ий слой соответствует глубинам от 0 до 10 м, II-ой слой охватывает глубины от 10 до 25 м и I-ый слой - от 25 м до дна. Каждый слой характеризуется своими условиями среды. Теоретически в каждом слое в любой момент времени могут находиться особи любого возраста каждой популяции (за некоторыми исключениями, о которых будет сказано ниже). Поэтому каждый ярус модельной системы должен содержать все элементы, которыми описывается модельное сообщество. Части популяций, находящиеся в данный момент времени в одном и том же слое, развиваются в соответствии с условиями среды данного слоя, вступают в трофические и конкурентные отношения между собой. Однако, если в соседнем слое условия среды, в первую очередь, температура воды, ближе к оптимальным для особей данной популяции, часть их может перейти в соседний слой. Перейдя в новый слой, такие особи продолжают развитие в соответствии с условиями этого слоя и подчиняются существующим в нем отношениям. В результате такого перехода состав зоопланктона каждого слоя меняется.
Особи каждой популяции делятся на несколько групп. Разбиение проводится по физиологическим, возрастным либо трофическим признакам, учет которых необходим для правильного описания развития сообщества. В популяции Bosmina obtusirostris lacustris Sars выделено 4 группы: ![]() - зимующие яйца (эфиппиумы), падающие на дно и дающие начало популяции весной (естественно, эта группа присутствует только в первом ярусе);
- зимующие яйца (эфиппиумы), падающие на дно и дающие начало популяции весной (естественно, эта группа присутствует только в первом ярусе); ![]() - партеногенетические яйца, развивающиеся в теле самки и обеспечивающие рост численности популяции летом;
- партеногенетические яйца, развивающиеся в теле самки и обеспечивающие рост численности популяции летом; ![]() - неполовозрелые особи;
- неполовозрелые особи; ![]() - половозрелые особи, способные к размножению. Спектр питания у представителей последних двух групп одинаков - это фитопланктон, бактерии и детрит. Остальные четыре популяции моделируемого сообщества (Eudiaptomus gracilis Sars, Mesocyclops oithonoides Sars, Limnocalanus macrurus Sars, Cyclops abyssorum Sars) относятся к веслоногим ракообразным. В каждой из них выделено пять групп. К первой группе (
- половозрелые особи, способные к размножению. Спектр питания у представителей последних двух групп одинаков - это фитопланктон, бактерии и детрит. Остальные четыре популяции моделируемого сообщества (Eudiaptomus gracilis Sars, Mesocyclops oithonoides Sars, Limnocalanus macrurus Sars, Cyclops abyssorum Sars) относятся к веслоногим ракообразным. В каждой из них выделено пять групп. К первой группе (![]() ,
, ![]() ,
, ![]() ,
, ![]() ) отнесены яйца, которые в популяциях E. gracilis, M. oithonoides и C. abyssorum развиваются в яйцевых мешках, а в популяции L. macrurus падают на дно и развиваются в соответствии с условиями придонного слоя. Поэтому группа яиц популяции L. macrurus (
) отнесены яйца, которые в популяциях E. gracilis, M. oithonoides и C. abyssorum развиваются в яйцевых мешках, а в популяции L. macrurus падают на дно и развиваются в соответствии с условиями придонного слоя. Поэтому группа яиц популяции L. macrurus (![]() ) присутствует только в первом ярусе модельного сообщества. Вторая группа особей популяций Copepoda (
) присутствует только в первом ярусе модельного сообщества. Вторая группа особей популяций Copepoda (![]() ,
, ![]() ,
, ![]() ,
, ![]() ) - группа науплиусов, в которую объединены особи всех науплиальных стадий развития данной популяции. Выделение их в отдельную группу необходимо, так как они резко отличаются от особей последующих копеподитных стадий морфологически, по средней массе тела, а в случае Cyclopoida и по спектру питания (Монаков, 1976). Питаются науплиусы всех популяций Copepoda фитопланктоном, бактериями и детритом, а сами могут служить пищей для хищных Cyclopoida. Третью группу особей популяций Copepoda составляют копеподиты I - III стадий развития (
) - группа науплиусов, в которую объединены особи всех науплиальных стадий развития данной популяции. Выделение их в отдельную группу необходимо, так как они резко отличаются от особей последующих копеподитных стадий морфологически, по средней массе тела, а в случае Cyclopoida и по спектру питания (Монаков, 1976). Питаются науплиусы всех популяций Copepoda фитопланктоном, бактериями и детритом, а сами могут служить пищей для хищных Cyclopoida. Третью группу особей популяций Copepoda составляют копеподиты I - III стадий развития (![]() ,
, ![]() ,
, ![]() ,
, ![]() ), которые сами питаются растительно-бактериальной пищей, но могут стать жертвой хищного зоопланктона. В четвертую группу (
), которые сами питаются растительно-бактериальной пищей, но могут стать жертвой хищного зоопланктона. В четвертую группу (![]() ,
, ![]() ,
, ![]() ,
, ![]() ) объединены особи IV - V копеподитных стадий, которые в популяциях Cyclopoida являются уже факультативными хищниками и добавляют к своему рациону мелких коловраток, инфузорий, особей популяции Cladocera, а также науплиусов и младших копеподитов Copepoda. И в последнюю, пятую, группу выделены половозрелые особи (
) объединены особи IV - V копеподитных стадий, которые в популяциях Cyclopoida являются уже факультативными хищниками и добавляют к своему рациону мелких коловраток, инфузорий, особей популяции Cladocera, а также науплиусов и младших копеподитов Copepoda. И в последнюю, пятую, группу выделены половозрелые особи (![]() ,
, ![]() ,
, ![]() ,
, ![]() ), которые отличаются от особей предыдущей группы тем, что они участвуют в размножении. Таким образом, всего имеется 24 группы зоопланктона, из которых 6 групп представляют собой яйца разных популяций.
), которые отличаются от особей предыдущей группы тем, что они участвуют в размножении. Таким образом, всего имеется 24 группы зоопланктона, из которых 6 групп представляют собой яйца разных популяций.
Кроме указанных общих свойств, некоторые из моделируемых популяций имеют в своем развитии особенности, которые также были учтены в модели. В частности, к таким особенностям относится явление диапаузы у особей Cyclopoida на определенной стадии их развития, выражающееся в том, что при наступлении неблагоприятных условий (обычно это бывает осенью) эти особи прекращают активную жизнедеятельность, падают на дно и возвращаются в активное состояние весной, обеспечивая возобновление популяции.
Как уже указывалось, особи каждой популяции, объединенные в одну группу, считались одинаковыми. Для каждой такой группы задавалась средняя масса особи и продолжительность развития в этой группе при оптимальной температуре и полном удовлетворении пищевых потребностей. Реальная скорость развития животных, то есть длительность их пребывания в данной группе, зависит от условий их обитания. В модели учитывались следующие факторы среды: температура места обитания, биомассы фитопланктона, детрита, мелких коловраток, простейших. Для определения смертности особей задавался также рацион рыб-планктофагов.
Как уже было сказано, изменение численности каждой группы зоопланктона данного слоя есть результат поступления особей в группу и убыли их в результате роста, размножения, гибели и сезонной вертикальной миграции (Гиляров, 1987). Рассмотрим, каким образом в модели описывается каждый их этих процессов.
Рост. Рост, то есть увеличение размера и массы организма, происходит в результате накопления энергии, поступающей в организм в процессе питания и расходующейся на энергетический обмен. В модели принято, что особи изменяют свою массу скачком при переходе из одной группы в другую (такое предположение оправдано тем, что все рассматриваемые виды относятся к классу ракообразных). Поэтому изменение скорости накопления энергии может проявляться только в изменении продолжительности пребывания особи на данной стадии развития, то есть в данной группе. Переход в следующую группу происходит в тот момент, когда накоплено достаточно много энергии для перехода.
На скорость энергетических процессов в первую очередь влияет температура. Влияние температуры среды на зоопланктон в модели описывалось при помощи температурных поправок. Для популяций теплолюбивого зоопланктона с оптимальной температурой ![]() °С была использована поправка И. В. Ивлевой (Ивлева, 1981), выражающаяся зависимостью
°С была использована поправка И. В. Ивлевой (Ивлева, 1981), выражающаяся зависимостью
![]() . (42)
. (42)
Для популяции L. macrurus, температурный оптимум которой принят равным ![]() °С, поправка
°С, поправка ![]() имела следующий вид:
имела следующий вид:
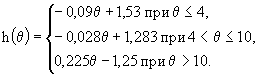 (43)
(43)
Для популяции C. abyssorum оптимальная температура была принята равной ![]() °С. Температурная поправка выражалась зависимостью
°С. Температурная поправка выражалась зависимостью
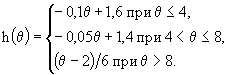 (44)
(44)
Если при оптимальной температуре особь i-ой группы имеет длительность развития ![]() , то при реальной температуре
, то при реальной температуре ![]() она будет развиваться медленнее, и продолжительность ее пребывания в этой группе будет равна
она будет развиваться медленнее, и продолжительность ее пребывания в этой группе будет равна
![]() . (45)
. (45)
Снизится и уровень энергетического обмена особи (![]() ), который в модели рассчитывается по ее весу
), который в модели рассчитывается по ее весу ![]() . С учетом температурной поправки, оксикалорийного коэффициента, калорийности зоопланктона (с), которая принималась равной 600 кал на 1 грамм сырого веса, и в пересчете на сутки формула трат особи на дыхание имела следующий вид:
. С учетом температурной поправки, оксикалорийного коэффициента, калорийности зоопланктона (с), которая принималась равной 600 кал на 1 грамм сырого веса, и в пересчете на сутки формула трат особи на дыхание имела следующий вид:
![]() (г/сутки) (46)
(г/сутки) (46)
Для Cladocera принималось ![]() 0,143,
0,143, ![]() 0,803; для Copepoda
0,803; для Copepoda ![]() 0,200,
0,200, ![]() 0,777 (Сущеня, 1972).
0,777 (Сущеня, 1972).
Кроме температуры, на скорость развития зоопланктона влияют и кормовые условия (очевидно, что здесь следует рассматривать только питающихся особей). Суточная потребность особи в пище (максимальный рацион) ![]() складывается из необходимости обеспечить ежесуточный весовой прирост и возместить энергию, затраченную на обмен:
складывается из необходимости обеспечить ежесуточный весовой прирост и возместить энергию, затраченную на обмен:
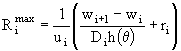 , (47)
, (47)
где ![]() ,
, ![]() - усвояемость пищи. Эти потребности должны быть удовлетворены за счет тех видов и того количества пищи, которые имеются в данный момент в распоряжении зоопланктона. Для мирного зоопланктона (все питающиеся группы, кроме
- усвояемость пищи. Эти потребности должны быть удовлетворены за счет тех видов и того количества пищи, которые имеются в данный момент в распоряжении зоопланктона. Для мирного зоопланктона (все питающиеся группы, кроме ![]() ,
, ![]() ,
, ![]() ,
, ![]() ) это суточные продукции фитопланктона (
) это суточные продукции фитопланктона (![]() ), бактерий (
), бактерий (![]() ), и биомасса детрита (
), и биомасса детрита (![]() ). Для факультативных хищников к этим видам пищи добавляются суточные продукции мелких коловраток (
). Для факультативных хищников к этим видам пищи добавляются суточные продукции мелких коловраток (![]() ), и простейших (
), и простейших (![]() ), а также биомассы зоопланктона модельных популяций: младших групп Copepoda
), а также биомассы зоопланктона модельных популяций: младших групп Copepoda
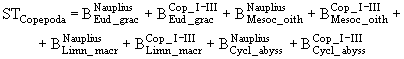 (48)
(48)
и особей B. obtusirostris ![]() . Известно, что животные разных видов и разного возраста не в одинаковой степени потребляют и усваивают разный корм. Этот факт отражался в модели заданием таблицы селективности питания S. Каждая строка такой таблицы задает распределение (в долях единицы) пищевых потребностей особи определенной группы по имеющимся видам пищи (в данном случае число видов пищи равно семи). Таким образом, сумма элементов каждой строки таблицы S равна единице. После этого могут быть определены суммарные потребности организмов i-ой группы в j-ом виде пищи:
. Известно, что животные разных видов и разного возраста не в одинаковой степени потребляют и усваивают разный корм. Этот факт отражался в модели заданием таблицы селективности питания S. Каждая строка такой таблицы задает распределение (в долях единицы) пищевых потребностей особи определенной группы по имеющимся видам пищи (в данном случае число видов пищи равно семи). Таким образом, сумма элементов каждой строки таблицы S равна единице. После этого могут быть определены суммарные потребности организмов i-ой группы в j-ом виде пищи:
![]() , (49)
, (49)
где ![]() соответствующие коэффициенты предпочтения организмами i-ой группы j-го вида пищи,
соответствующие коэффициенты предпочтения организмами i-ой группы j-го вида пищи, ![]() - численность i-ой группы. Кроме хищного зоопланктона, пищей
- численность i-ой группы. Кроме хищного зоопланктона, пищей ![]() и
и ![]() питаются рыбы. Рацион рыб FishT, приходящийся за единицу времени на долю модельного зоопланктона, делится между Cladocera и Copepoda в долях
питаются рыбы. Рацион рыб FishT, приходящийся за единицу времени на долю модельного зоопланктона, делится между Cladocera и Copepoda в долях ![]() и
и ![]() соответственно. Поэтому общая потребность в Cladocera, то есть в особях модельной популяции B. obtusirostris, оказывается равной
соответственно. Поэтому общая потребность в Cladocera, то есть в особях модельной популяции B. obtusirostris, оказывается равной
![]() , (50)
, (50)
а в младших группах Copepoda -
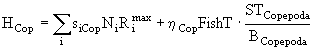 , (51)
, (51)
где ![]() - суммарная биомасса всех модельных Copepoda.
- суммарная биомасса всех модельных Copepoda.
Реальная обеспеченность зоопланктона пищей рассчитывалась по схеме В. С. Ивлева (Ивлев, 1955) следующим образом. Зная имеющееся количество каждого вида пищи ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() (для краткости будем обозначать
(для краткости будем обозначать ![]() ) и потребность в нем
) и потребность в нем ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() (для краткости будем обозначать
(для краткости будем обозначать ![]() ), можно определить для него так называемую напряженность пищевых отношений
), можно определить для него так называемую напряженность пищевых отношений
![]() , (52)
, (52)
которая служит основой для расчета степени удовлетворения общей потребности в этом виде пищи:
![]() . (53)
. (53)
Так как особи i-ой группы зоопланктона удовлетворяют за счет пищи ![]()
![]() -ю часть своего рациона, то, зная максимальные рационы, можно рассчитать реальные рационы:
-ю часть своего рациона, то, зная максимальные рационы, можно рассчитать реальные рационы: ![]() и соответствующие степени удовлетворения пищевых потребностей
и соответствующие степени удовлетворения пищевых потребностей
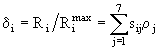 . (54)
. (54)
Влияние степени удовлетворения пищевых потребностей особи на продолжительность ее пребывания ![]() в i-ой группе при данной температуре задавалось формулой, основанной на данных по питанию планктонных ракообразных фильтраторов (Weglenska, 1968):
в i-ой группе при данной температуре задавалось формулой, основанной на данных по питанию планктонных ракообразных фильтраторов (Weglenska, 1968):
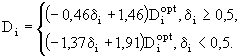 (55)
(55)
Если особь при данных условиях среды должна развиваться в i-ой группе ![]() суток, то за одни сутки она успевает пройти
суток, то за одни сутки она успевает пройти ![]() часть необходимого развития. В следующие сутки условия среды могут измениться, и соответственно изменится скорость развития. Для особи, пришедшей в i-ую группу в момент времени
часть необходимого развития. В следующие сутки условия среды могут измениться, и соответственно изменится скорость развития. Для особи, пришедшей в i-ую группу в момент времени ![]() , моментом перехода в следующую группу считалось то время Т, для которого оказывается
, моментом перехода в следующую группу считалось то время Т, для которого оказывается
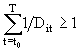 . (56)
. (56)
Таким образом, реальное пребывание особи в данной группе, то есть реальная скорость развития, оказывается величиной переменной, зависящей от меняющихся условий среды. Для особей старших групп каждой популяции момент завершения развития в группе интерпретировался как момент их естественной гибели.
Гибель особей. Кроме естественной, в модели учитывались еще две причины гибели особей: гибель при голодании и выедание хищниками.
Предполагалось, что гибель от голода может наступить тогда, когда степень удовлетворения пищевых потребностей особи ![]() меньше того критического значения
меньше того критического значения ![]() , при котором потребленной пищи хватает только для возмещения энергетических затрат на обмен. Определив критическую степень удовлетворения пищевых потребностей
, при котором потребленной пищи хватает только для возмещения энергетических затрат на обмен. Определив критическую степень удовлетворения пищевых потребностей
![]() , (57)
, (57)
можно считать, что коэффициент смертности от голода равен
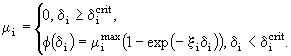 (58)
(58)
Если ![]() - часть рациона хищников, приходящаяся на особей i-ой группы, а
- часть рациона хищников, приходящаяся на особей i-ой группы, а ![]() - биомасса этой группы, то коэффициент смертности от воздействия хищников при отсутствии других причин гибели можно представить, например, в виде
- биомасса этой группы, то коэффициент смертности от воздействия хищников при отсутствии других причин гибели можно представить, например, в виде
![]() . (59)
. (59)
Формула (59) учитывает зависимость степени выедания зоопланктона как от количества самого зоопланктона, так и от рациона хищников и отражает тот факт, что с увеличением плотности жертв или рациона хищников смертность возрастает и стремится к какому-то предельному значению ![]() , благодаря чему полное выедание жертвы невозможно (Ивлев, 1955). Для групп
, благодаря чему полное выедание жертвы невозможно (Ивлев, 1955). Для групп ![]() ,
, ![]() величины
величины ![]() и
и ![]() определяются следующим образом:
определяются следующим образом:
![]() ,
, ![]() . (60)
. (60)
Для групп младших Copepoda ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() величины
величины ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() задаются соотношениями
задаются соотношениями
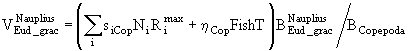 ,
,
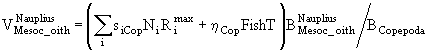 ,
,
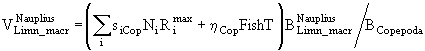 ,
,
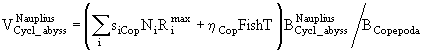 ,
,
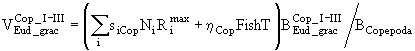 ,
,
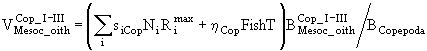 ,
,
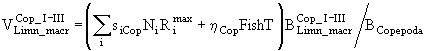 ,
,
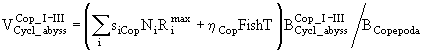 . (61)
. (61)
Для старших Copepoda ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() , для которых хищниками являются только рыбы, величины
, для которых хищниками являются только рыбы, величины ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() определяются следующим образом:
определяются следующим образом:
![]() ,
,
![]() ,
,
![]() ,
,
![]() ,
,
![]() ,
,
![]() ,
,
![]() ,
,
![]() . (62)
. (62)
Коэффициент общей смертности определяется по формуле
![]() . (63)
. (63)
Для тех популяций, в которых яйца вынашиваются самками, смертность таких яиц считалась равной смертности взрослых. Для яиц L. macrurus по данным (Roff, 1972) была построена зависимость выживаемости от температуры за пять суток (шаг модели равен пяти суткам):
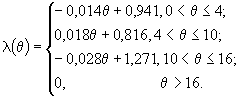 (64)
(64)
Смертность яиц лимнокалянуса считалась равной
![]() . (65)
. (65)
Процент гибели эфиппиумов популяции B. obtusirostris принимался равным 80%.
Размножение. Процесс размножения описывается по единой схеме для всех популяций, кроме L. macrurus. Размножение особей и тем самым пополнение первой группы вычисляется по количеству яиц ![]() , отложенных в популяции за единицу времени:
, отложенных в популяции за единицу времени:
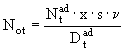 , (66)
, (66)
где ![]() ,
, ![]() - соответственно число и продолжительность развития половозрелых особей, x - соотношение полов (
- соответственно число и продолжительность развития половозрелых особей, x - соотношение полов (![]() ), s - среднее число кладок у самки в течение ее жизни (за репродуктивный период),
), s - среднее число кладок у самки в течение ее жизни (за репродуктивный период), ![]() - количество яиц в кладке в расчете на половозрелую самку
- количество яиц в кладке в расчете на половозрелую самку ![]() , то есть плодовитость самки. Формула (66) не совсем удобна, поскольку в ней необходимо задавать довольно изменчивую и недостаточно изученную величину соотношения полов в популяции зоопланктона. Гораздо более предпочтительным оказывается использование другого параметра - плодовитости в расчете на половозрелую особь. Если принять во внимание, что
, то есть плодовитость самки. Формула (66) не совсем удобна, поскольку в ней необходимо задавать довольно изменчивую и недостаточно изученную величину соотношения полов в популяции зоопланктона. Гораздо более предпочтительным оказывается использование другого параметра - плодовитости в расчете на половозрелую особь. Если принять во внимание, что ![]() , а значит
, а значит ![]() , то формула (66) принимает следующий вид
, то формула (66) принимает следующий вид
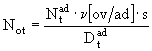 . (67)
. (67)
Плодовитости ![]() в расчете на половозрелую особь для каждой из указанных популяций задавались в данном варианте модели два раза в месяц с линейной интерполяцией для промежуточных моментов времени. Ювенильные особи модельной популяции B. obtusirostris, появившиеся из эфиппиумов, в момент появления оказываются в придонном слое.
в расчете на половозрелую особь для каждой из указанных популяций задавались в данном варианте модели два раза в месяц с линейной интерполяцией для промежуточных моментов времени. Ювенильные особи модельной популяции B. obtusirostris, появившиеся из эфиппиумов, в момент появления оказываются в придонном слое.
Процесс размножения в популяции L. macrurus имеет свои особенности. Например, по наблюдениям на оз. Врево (Авинский, Лихарева, 1986) у L. macrurus, после завершения метаморфоза, то есть по достижении половозрелости, откладка яиц у самок начинается не сразу, а после шестимесячной паузы, сопровождаемой снижением численности популяции. Затем начинается размножение, и самка делает в среднем три кладки. Закончив размножение, самка погибает. В модели этот процесс был описан следующим образом. Степень развития, достигнутая взрослой особью к данному моменту времени t, определяется как 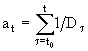 , где
, где ![]() - время поступления особи в группу взрослых,
- время поступления особи в группу взрослых, ![]() - продолжительность развития особи в этой группе при условиях среды, наблюдающихся в момент времени
- продолжительность развития особи в этой группе при условиях среды, наблюдающихся в момент времени ![]() . Было принято, что особь может начинать размножение только после того, как достигнет уровня развития
. Было принято, что особь может начинать размножение только после того, как достигнет уровня развития ![]() . Первую кладку совершают особи, для которых
. Первую кладку совершают особи, для которых ![]() . Вторую кладку могут производить особи, для которых
. Вторую кладку могут производить особи, для которых ![]() , и третью - те, для которых
, и третью - те, для которых ![]() . При этом
. При этом ![]() . По достижении
. По достижении ![]() особь погибает. Количество яиц
особь погибает. Количество яиц ![]() и доля самок соответствующего возраста со сперматофорами (то есть способных отложить яйца)
и доля самок соответствующего возраста со сперматофорами (то есть способных отложить яйца) ![]() задавались для каждой k-ой кладки. Таким образом, число отложенных в популяции яиц в момент времени t равно
задавались для каждой k-ой кладки. Таким образом, число отложенных в популяции яиц в момент времени t равно
![]() , (68)
, (68)
где ![]() - число взрослых особей, достигших возраста k-ой кладки. Яйца откладывают самки, находящиеся в любом слое, но все отложенные яйца падают на дно, то есть в первых слой, и там происходит их дальнейшее развитие.
- число взрослых особей, достигших возраста k-ой кладки. Яйца откладывают самки, находящиеся в любом слое, но все отложенные яйца падают на дно, то есть в первых слой, и там происходит их дальнейшее развитие.
Диапауза в развитии циклопов. Явление диапаузы в развитии циклопов разных видов очень сложно и разнообразно в своем проявлении. В разных популяциях диапауза может наступать как в разное время года, так и на разных стадиях развития особей. О диапаузе в популяциях Cyclopidae Ладожского озера практически нет никаких достоверных данных. Поэтому необходимые предположения были сделаны частично на основе литературных данных, а частично из соображений успешности моделирования.
Как для популяции M. oithonoides, так и для C. abyssorum было принято, что диапауза наступает на IV-V копеподитных стадиях (группы ![]() ,
, ![]() ). В действительности уход особей в диапаузу и выход из нее растянуты во времени и определяются физиологическим состоянием особи. Однако при моделировании мы вынуждены приурочить их к определенным моментам времени, которые задавались для каждой популяции отдельно. Считалось, что в момент наступления диапаузы уходит из популяции вся группа старших копеподитов, причем во время пребывания в диапаузе особи этой группы сохраняют тот уровень развития, которого они сумели достичь к этому моменту времени. Начиная с этого момента, младшие копеподиты, которые завершили развитие в своей группе и должны перейти в следующую, сразу уходят в диапаузу, имея нулевую степень развития в группе старших копеподитов. В момент выхода из диапаузы выходит в первый слой вся группа старших копеподитов, особи которой продолжают свое развитие с того состояния, в котором они находились в период диапаузы. Таким образом, репродуктивного состояния они достигают в разное время, что обеспечивает растянутость периода восстановления популяции после диапаузы. Было сделано предположение о том, что в период диапаузы гибнет 0,1 часть особей, пребывающих в этом состоянии.
). В действительности уход особей в диапаузу и выход из нее растянуты во времени и определяются физиологическим состоянием особи. Однако при моделировании мы вынуждены приурочить их к определенным моментам времени, которые задавались для каждой популяции отдельно. Считалось, что в момент наступления диапаузы уходит из популяции вся группа старших копеподитов, причем во время пребывания в диапаузе особи этой группы сохраняют тот уровень развития, которого они сумели достичь к этому моменту времени. Начиная с этого момента, младшие копеподиты, которые завершили развитие в своей группе и должны перейти в следующую, сразу уходят в диапаузу, имея нулевую степень развития в группе старших копеподитов. В момент выхода из диапаузы выходит в первый слой вся группа старших копеподитов, особи которой продолжают свое развитие с того состояния, в котором они находились в период диапаузы. Таким образом, репродуктивного состояния они достигают в разное время, что обеспечивает растянутость периода восстановления популяции после диапаузы. Было сделано предположение о том, что в период диапаузы гибнет 0,1 часть особей, пребывающих в этом состоянии.
Вертикальные миграции. Все процессы, которые были описаны, являются основными в развитии популяций и происходят во всех трех слоях водной толщи, в каждом слое при своих условиях и со своей интенсивностью. Однако изменение численности популяций в каждом слое происходит не только в результате размножения, роста и гибели особей, но и перехода их из слоя в слой в поисках более благоприятных условий (Рудяков, 1986). Считалось, что особь может в данный момент перейти в соседний слой, выше- или нижележащий. При определении стратегии перехода в первую очередь принимались во внимание температурные условия.
Для особей каждой популяции, находящихся в соседних слоях, сравнивались температурные поправки на данный момент времени. Чем ближе значение поправки к единице, тем ближе температура к оптимальной (очевидно, что все значения поправок, в том числе и поправки Ивлевой для ![]() °С, не меньше единицы). Особь стремится перейти из слоя с большим значением поправки (дальше от единицы, то есть от оптимальных условий) в слой с меньшим значением. Если температуры, а следовательно, и значения поправок в соседних слоях примерно одинаковы, необходимость перехода определяется пищевыми условиями: особи идут туда, где степени удовлетворения пищевых потребностей особей данной группы выше. По разности соответствующих показателей (А) вычислялась часть особей данной группы (s), которая должна перейти в соседний слой:
°С, не меньше единицы). Особь стремится перейти из слоя с большим значением поправки (дальше от единицы, то есть от оптимальных условий) в слой с меньшим значением. Если температуры, а следовательно, и значения поправок в соседних слоях примерно одинаковы, необходимость перехода определяется пищевыми условиями: особи идут туда, где степени удовлетворения пищевых потребностей особей данной группы выше. По разности соответствующих показателей (А) вычислялась часть особей данной группы (s), которая должна перейти в соседний слой:
![]() . (69)
. (69)
Перешедшие особи сохраняли достигнутую к моменту перехода степень своего развития и продолжали развиваться в новых условиях.
Вычисление продукционных характеристик популяций. Описывая динамику численности возрастных групп популяций во всех вертикальных слоях, модель дает возможность рассчитать биомассы, продукции, элиминации каждой популяции за любой период времени. Так, если ![]() - численность i-ой группы зоопланктона в момент времени t,
- численность i-ой группы зоопланктона в момент времени t, ![]() - масса особи этой группы, то биомасса группы в момент времени t равна
- масса особи этой группы, то биомасса группы в момент времени t равна
![]() . (70)
. (70)
Продукция группы за какой-то период времени есть прирост биомассы группы за это время. В модели продукцией ![]() , создаваемой i-ой группой за один шаг модельного времени
, создаваемой i-ой группой за один шаг модельного времени ![]() считалось увеличение биомассы, получающееся при переходе
считалось увеличение биомассы, получающееся при переходе ![]() особей из i-ой группы в следующую, i + 1-ю, группу:
особей из i-ой группы в следующую, i + 1-ю, группу:
![]() . (71)
. (71)
Если i-ая группа является последней для популяции, то выбывающие из не в процессе роста особи гибнут и тем самым изымают из группы соответствующую биомассу, которая составляет часть элиминации ![]() группы. Другую часть элиминации групп половозрелых особей образует биомасса особей, гибнущих по другим причинам. Таким образом, для
группы. Другую часть элиминации групп половозрелых особей образует биомасса особей, гибнущих по другим причинам. Таким образом, для ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() элиминация определяется соотношением
элиминация определяется соотношением
![]() , (72)
, (72)
где ![]() - смертность особей i-ой группы от хищника и голодания. Для групп неполовозрелых особей элиминация определяется только их смертностью и находится по формуле:
- смертность особей i-ой группы от хищника и голодания. Для групп неполовозрелых особей элиминация определяется только их смертностью и находится по формуле:
![]() . (73)
. (73)
Биомасса, продукция и элиминация популяций мирного зоопланктона определяется суммированием соответствующих показателей для отдельных возрастных групп данной популяции. Исключение составляют величины продукций факультативных хищников M. oithonoides и C. abyssorum, так как часть продукции науплиусов и младших копеподитов этих популяций используется в пищу старшими особями своих же популяций и при суммировании учитывалась бы дважды. Поэтому от суммарной продукции всех возрастных групп для этих популяций надо вычесть продукцию младших особей, съеденную старшими.
Зная продукционные показатели в каждый момент времени, легко найти среднюю биомассу, а также продукцию и элиминацию популяций для любого интервала времени - месяца, сезона, года. Суммирование соответствующего показателя популяции по всем слоям даст его значение для всего столба воды. Соответствующим суммированием определяются и характеристики групп мирного и хищного зоопланктона.
Некоторые физиологические характеристики особей и параметры модели. Средние массы для большинства групп рассчитаны по формулам вида ![]() , где l (мм) - средняя длина особи i-ой группы, определенная по данным проб,
, где l (мм) - средняя длина особи i-ой группы, определенная по данным проб, ![]() и
и ![]() - параметры, рекомендуемые для расчета массы особи k-го вида. Массы яиц и науплиусов задавались по другим соображениям или брались непосредственно из литературы. Средние массы особей по всем группам зоопланктона представлены в таблице 1.
- параметры, рекомендуемые для расчета массы особи k-го вида. Массы яиц и науплиусов задавались по другим соображениям или брались непосредственно из литературы. Средние массы особей по всем группам зоопланктона представлены в таблице 1.
Таблица 1. Средние массы тела ![]() (мг) особей i-ой группы моделируемых популяций.
(мг) особей i-ой группы моделируемых популяций.
|
Группа |
B. obt. |
E. grac. |
M. oith. |
L. macr. |
C. abys. |
|
1 |
0.0008 |
0.0007 |
0.0003 |
0.0005 |
0.0005 |
|
2 |
0.0008 |
0.0030 |
0.0005 |
0.0041 |
0.0030 |
|
3 |
0.0095 |
0.0047 |
0.0011 |
0.0066 |
0.0156 |
|
4 |
0.0256 |
0.0200 |
0.0055 |
0.0411 |
0.0418 |
|
5 |
- |
0.0517 |
0.0108 |
0.1940 |
0.0892 |
|
|
0.0500 |
0.0543 |
0.0134 |
0.2039 |
0.0936 |
![]() - так называемая "дефинитивная масса", которой якобы стремится достичь взрослое животное. Введение этой величины необходимо для расчета пищевых потребностей взрослых особей.
- так называемая "дефинитивная масса", которой якобы стремится достичь взрослое животное. Введение этой величины необходимо для расчета пищевых потребностей взрослых особей.
Продолжительность развития особей в каждой группе при оптимальной температуре и полном удовлетворении пищевых потребностей (![]() ) первоначально задавались на основе литературных данных и некоторых экспериментальных данных по зоопланктону Ладожского озера. Так как именно скорость развития особей определяет моменты перехода их из стадии в стадию, то есть динамику популяции, то при отладке модели величины
) первоначально задавались на основе литературных данных и некоторых экспериментальных данных по зоопланктону Ладожского озера. Так как именно скорость развития особей определяет моменты перехода их из стадии в стадию, то есть динамику популяции, то при отладке модели величины ![]() корректировались таким образом, чтобы обеспечить желаемую динамику данных конкретных популяций. Работа с моделью показала, что именно к изменениям параметра
корректировались таким образом, чтобы обеспечить желаемую динамику данных конкретных популяций. Работа с моделью показала, что именно к изменениям параметра ![]() она наиболее чувствительна. В конечном итоге были приняты следующие значения
она наиболее чувствительна. В конечном итоге были приняты следующие значения ![]() (табл. 2).
(табл. 2).
Таблица 2. Продолжительности развития![]() (сут) особей i-ой группы при оптимальной температуре и полном удовлетворении пищевых потребностей.
(сут) особей i-ой группы при оптимальной температуре и полном удовлетворении пищевых потребностей.
|
Группа |
B. obt. |
E. grac. |
M. oith. |
L. macr. |
C. abys. |
|
1 |
15,00 |
1,50 |
2,30 |
10,00 |
4,00 |
|
2 |
1,25 |
3,00 |
7,00 |
75,00 |
15,00 |
|
3 |
2,50 |
2,00 |
6,50 |
7,50 |
10,00 |
|
4 |
7,50 |
1,50 |
6,75 |
5,00 |
10,00 |
|
5 |
- |
15,00 |
7,50 |
101,50 |
32,50 |
Основой для задания плодовитостей в расчете на половозрелую особь для всех популяций, кроме L. macrurus, служили данные наблюдений, однако при отладке модели они изменялись таким образом, чтобы обеспечить нужное абсолютное количество зоопланктона. Значения плодовитостей являются вторыми по значимости при оценке чувствительности модели к изменению ее параметров. Принятые при отладке модели значения плодовитости приведены в таблице 3.
Таблица 3. Принятые в модели значения плодовитости в расчете на половозрелую особь.
|
Месяц |
B. obt. |
E. grac. |
M. oith. |
L. macr. |
C. abys. |
|
VII |
0 |
1,50 |
1,00 |
3,00 |
0,50 |
|
|
0 |
1,20 |
1,20 |
2,00 |
1,00 |
|
VIII |
0,15 |
0,80 |
2,00 |
1,00 |
1,50 |
|
|
0,30 |
0,30 |
1,50 |
0,50 |
0,80 |
|
IX |
0,40 |
0,10 |
0,80 |
0,40 |
0,50 |
|
|
0,50 |
0 |
0,60 |
0,30 |
0,25 |
|
X |
0,20 |
0 |
0,30 |
0,20 |
0 |
|
|
0,05 |
0 |
0,20 |
0,10 |
0 |
|
XI |
0 |
0 |
0,15 |
0 |
0 |
|
|
0 |
0 |
0,10 |
0 |
0 |
|
XII |
0 |
0 |
0,05 |
0 |
0 |
|
|
0 |
0 |
0,05 |
0 |
0 |
|
I |
0 |
0 |
0,05 |
0 |
0 |
|
|
0 |
0 |
0,05 |
0 |
0 |
|
II |
0 |
0 |
0,10 |
0 |
0 |
|
|
0 |
0 |
1,00 |
0 |
0 |
|
III |
0 |
0 |
3,00 |
0 |
0 |
|
|
0 |
0 |
5,00 |
0 |
0 |
|
IV |
0 |
0 |
7,00 |
0 |
0 |
|
|
0 |
0 |
2,50 |
0 |
0 |
|
V |
0 |
0 |
0,50 |
0 |
0 |
|
|
0 |
1,75 |
0,30 |
10,00 |
1,00 |
|
VI |
0 |
1,40 |
0,25 |
7,50 |
2,00 |
|
|
0 |
1,50 |
0,20 |
5,00 |
1,00 |
|
Число кладок |
- |
4 |
5 |
3 |
4 |
Для популяции L. macrurus принималось, что самка делает 3 кладки размером 70, 50 и 30 яиц соответственно, причем самки со сперматофорами составляют 70% среди особей, достигших репродуктивного возраста. При этом, если все развитие особи в стадии взрослых принять за 1, то первая кладка производится, когда самка находится "в возрасте" от 0,875 до 0,885, вторая - от 0,895 до 0,905 и третья - от 0,915 до 0,925.
При описании явления диапаузы в развитии моделируемых популяций Cyclopidae было принято, что старшие копеподиты популяции M. oithonoides уходят в диапаузу 15 ноября, а популяции C. abyssorum -30 октября. Выход из диапаузы для обеих популяций задавался 18 мая.
Матрица селективности питания групп зоопланктона каждым видом пищи приведена в таблице 4.
Таблица 4. Селективность питания модельного зоопланктона.
| Группа | |||||||
| 0,50 | 0,50 | 0 | 0 | 0 | 0 | 0 | |
| 0,50 | 0,50 | 0 | 0 | 0 | 0 | 0 | |
| 0,60 | 0 | 0 | 0,40 | 0 | 0 | 0 | |
| 0,75 | 0,15 | 0 | 0,10 | 0 | 0 | 0 | |
| 0,75 | 0,15 | 0 | 0,10 | 0 | 0 | 0 | |
| 0,75 | 0,15 | 0 | 0,10 | 0 | 0 | 0 | |
| 0,40 | 0,15 | 0 | 0,10 | 0,35 | 0 | 0 | |
| 0,40 | 0,15 | 0 | 0,10 | 0,35 | 0 | 0 | |
| 0,03 | 0,02 | 0,15 | 0,20 | 0,30 | 0,20 | 0,10 | |
| 0,03 | 0,02 | 0,15 | 0,20 | 0,30 | 0,20 | 0,10 | |
| 0,70 | 0,10 | 0 | 0,10 | 0,10 | 0 | 0 | |
| 0,70 | 0,10 | 0 | 0,10 | 0,10 | 0 | 0 | |
| 0,80 | 0,05 | 0 | 0,05 | 0,05 | 0,05 | 0 | |
| 0,80 | 0,05 | 0 | 0,05 | 0,05 | 0,05 | 0 | |
| 0,40 | 0,15 | 0 | 0,10 | 0,35 | 0 | 0 | |
| 0,40 | 0,15 | 0 | 0,10 | 0,35 | 0 | 0 | |
| 0,03 | 0,02 | 0,15 | 0,20 | 0,30 | 0,20 | 0,10 | |
| 0,03 | 0,02 | 0,15 | 0,20 | 0,30 | 0,20 | 0,10 |
В модели принято, что максимально возможная смертность ![]() для всех групп одинакова и равна 0,3. Рацион рыб-планктофагов делился между Cladocera и Copepoda в долях
для всех групп одинакова и равна 0,3. Рацион рыб-планктофагов делился между Cladocera и Copepoda в долях ![]() и
и ![]() .
.
Дополнительная информация:
© 2001-2026 Кафедра биофизики МГУ