Известно, что вертикальное и горизонтальное пространственное распределение планктона в водной среде отличается высокой неоднородностью. Пятнистость ("пэтчинг") распределения планктона в океанах, морях, больших и малых озерах обнаруживается в очень широком диапазоне пространственных и временных масштабов. Практически все современные исследования неоднородного распределения планктона относятся к большим водным системам: океанам, морям, крупным озерам и водохранилищам. Данные наблюдений свидетельствуют о том, что на пространственных масштабах от десяти километров и более пятнистость планктонного распределения определяется в основном неоднородностью соответствующих физико-химических полей (Malchow et al., 2002). Для фитопланктона часто можно установить прямое соответствие между неоднородностью его распределения и физическими (абиотическими) факторами среды как по вертикали (явления в термоклине, конвективный перенос, апвеллинг) (Fennel, 1999), так и по горизонтали (воздействие ветра, фронты и синоптические вихри) (Webster, 1990; Verhagen, 1994; Webster, Hutchinson, 1994; Kimura et al., 1997; Fennel, 2001). Причины пространственной неоднородности и изменчивости зоопланктона менее ясны, поскольку они способны самостоятельно совершать достаточно значительные вертикальные и горизонтальные перемещения, а не только пассивно переноситься движением воды. На масштабах менее 100 метров определяющим является влияние турбулентного обмена. Следует иметь в виду, что большие амплитуды изменчивости планктона на мезомасштабе (1 - 100 км) часто наблюдаются и в отсутствие значительных физических воздействий на него. В этих случаях уже трудно найти простое соответствие между пространственными распределениями температуры, хлорофилла и биомассой планктона, хотя иногда удается установить слабые отрицательные корреляции между фито- и зоопланктоном (Mackas, Boyd, 1979; Steele, Henderson, 1979). Самым сложным является промежуточный диапазон от 100 метров до 10 км, где нет однозначной зависимости между пятнистостью и каким-то одним определяющим фактором. В пределах этого диапазона зависимость, как правило, многофакторная и при прочих равных условиях с примерно одинаковым вкладом каждого из факторов. Можно также говорить о том, что по всей видимости превалирующим здесь является биологический, а не гидродинамический механизм формирования пятнистости планктона (Malchow et al., 2002). При относительно однородных внешних физических условиях важно рассмотреть, действительно ли пятнистость распределения представляет собой следствие экологических процессов, а не просто чисто внешних воздействий на систему.
Таким образом, мы вновь сталкиваемся с проблемой выделения характеристической длины и временных масштабов. Известно, что изменчивость различных процессов и явлений в океане охватывает широкий диапазон пространственных и временных масштабов. Весь этот широкий спектр изменчивости можно рассматривать как множество более узких спектральных диапазонов (иногда перекрывающихся), в пределах каждого из которых выделяется некоторый доминантный фактор, управляющий движением жидкости и влияющий на биологические процессы. По всей видимости можно говорить о том, что абиотические факторы задают определенную иерархию внешних воздействий, которые, в свою очередь, определяют разнообразные механизмы биологического ответа на различных временных и пространственных масштабах (Mackas et al., 1985).
Термин "пятнистость" обычно используется для описания изменчивости на горизонтальных масштабах от 10 м до 100 км и на вертикальных масштабах от 0,1 м до 50 м. С этими диапазонами связывают несколько физических режимов (Bennett, Denman, 1985; Abraham, 1998). Примерно между 1 мм и 0,1 м (этот диапазон меньше того диапазона, где мы без труда можем обнаружить изменчивость концентрации организмов) движение жидкости можно считать изотропным. Сглаживание возникающих пространственных градиентов за счет молекулярной диффузии и вязкости происходит достаточно быстро и эффективно. Эту область спектра изменчивости называют вязким поддиапазоном или диапазоном диссипации кинетической энергии в тепло. На несколько больших масштабах, примерно от 0,1 м до 1 м (эту область называют диапазоном равновесия или инерционным диапазоном), - флуктуации скорости продолжают оставаться изотропными. В теории течений считается, что в пределах этого спектрального диапазона приток или отток кинетической энергии достаточно незначителен. Энергия переносится взаимодействующими турбулентными вихрями на более мелкие масштабы, где она фактически диссипирует. На масштабах больше нескольких метров (диапазон плавучести), флуктуации вертикальной скорости в значительной степени подавляются за счет устойчивой стратификации плотности, а турбулентные и периодические движения становятся все в большей степени горизонтальными. Наименьший предел для этого диапазона устанавливается в соответствии со степенью стратификации и измеряется с помощью частоты плавучести ![]() (частоты Брента-Вяйсяля) и масштаба длины Озмидова
(частоты Брента-Вяйсяля) и масштаба длины Озмидова ![]() , где g - ускорение свободного падения,
, где g - ускорение свободного падения, ![]() - плотность воды,
- плотность воды, ![]() - скорость диссипации турбулентной кинетической энергии. Там, где вертикальные градиенты плотности в верхнем слое незначительны, флуктуации скорости могут быть практически изотропными во всем перемешанном слое (до 10 м и даже более). Верхний предел горизонтального масштаба практически равен локальному внутреннему радиусу деформации Россби:
- скорость диссипации турбулентной кинетической энергии. Там, где вертикальные градиенты плотности в верхнем слое незначительны, флуктуации скорости могут быть практически изотропными во всем перемешанном слое (до 10 м и даже более). Верхний предел горизонтального масштаба практически равен локальному внутреннему радиусу деформации Россби: ![]() , где h - локальная толщина водного столба, f - параметр Кориолиса. Этот масштаб длины имеет порядок 10 - 20 км и важен в тех случаях, когда необходимо учитывать ускорение за счет действия силы Кориолиса. В пределах диапазона плавучести одним из наиболее важных видов движений являются внутренние волны. На еще больших горизонтальных масштабах (примерно от 10 до 1000 км) поток в пределах верхнего слоя океана является по существу геострофическим и двумерным. По всей видимости, в первом приближении можно полагать, что характерные временные масштабы для развития и сохранения динамических структур разного типа в пределах каждой из этих спектральных областей пропорциональны соответствующим горизонтальным масштабам длины. Перераспределение колебаний между масштабами формирует направленный турбулентный каскад переноса кинетической энергии от более крупных образований к более мелким, вплоть до ее диссипации на малых масштабах.
, где h - локальная толщина водного столба, f - параметр Кориолиса. Этот масштаб длины имеет порядок 10 - 20 км и важен в тех случаях, когда необходимо учитывать ускорение за счет действия силы Кориолиса. В пределах диапазона плавучести одним из наиболее важных видов движений являются внутренние волны. На еще больших горизонтальных масштабах (примерно от 10 до 1000 км) поток в пределах верхнего слоя океана является по существу геострофическим и двумерным. По всей видимости, в первом приближении можно полагать, что характерные временные масштабы для развития и сохранения динамических структур разного типа в пределах каждой из этих спектральных областей пропорциональны соответствующим горизонтальным масштабам длины. Перераспределение колебаний между масштабами формирует направленный турбулентный каскад переноса кинетической энергии от более крупных образований к более мелким, вплоть до ее диссипации на малых масштабах.
Турбулентный перенос оказывает основное влияние на образование пятен планктона и именно его рассматривают как один из важных источников биологической изменчивости. В первоначальной KISS-модели (KIerstead - Slobodkin - Skellam) (Skellam, 1951; Kierstead, Slobodkin, 1953; Okubo, 1980) было показано, что локализованное цветение популяции, имеющей высокую скорость роста, будет иметь место только в том случае, если пространственные масштабы этого образования достаточно велики для того, чтобы рост популяции превышал ее рассеяние из пятна вследствие процессов адвекции и турбулентности. KISS-модель параметризует процесс турбулентного перемешивания введением коэффициента турбулентного обмена. При этом не учитывается мелкомасштабная изменчивость в турбулентном каскаде. Другие модели (Denman, Platt, 1976; Denman et al., 1977; Wilson et al., 1979) пытаются избежать этого ограничения с помощью предсказания формы спектра фитопланктонной изменчивости. Подобно KISS-модели, в них определяется характеристический масштаб длины, на котором рост популяции планктона становится настолько же важен, как и процесс турбулентного разрушения пятна. Следует отметить еще и тот факт, что все указанные модели основаны на предположении об изотропности турбулентности (которое для океанских условий не выполняется уже на масштабах, больших нескольких метров) и используют одинаковую скорость роста популяции в любой точке пространства. Между тем, имеются данные наблюдений, показывающие, что удельные скорости роста природных популяций фитопланктона и веслоногих ракообразных имеют вполне определенную собственную пространственно-временную структуру, хотя у популяции зоопланктона размах изменчивости несколько меньше (Therriault et al., 1978; Kimmerer, 1983).
Еще одним возможным источником возникновения пространственных неоднородностей являются чисто биологические взаимодействия между трофическими уровнями или конкурентные отношения между видами, которые обычно моделируются системами нелинейных дифференциальных или разностных уравнений. Включение в такого рода системы диффузии (в некоторых моделях и адвекции) может приводить к неустойчивости и образованию в конечном итоге пространственно-временных неоднородностей. В ряде случаев образование зон повышенной концентрации биогенных веществ, планктона, рыб может быть связано с процессами апвеллинга или вертикального перемешивания, с образованием фронтальных зон, вихревых образований различной физической природы, приливами (Kimura et al., 1997; Fennel, 1999; Fennel, 2001).
Одним из широко используемых методов описания пространственной изменчивости является спектральное разложение Фурье в зависимости от волновых чисел. Первоначально этот метод был использован для анализа полей распределения хлорофилла. При этом было показано, что спектр хлорофилла близок к наблюдаемым спектрам температуры. На этом основании был сделан вывод о том, что пятнистость распределения фитопланктона, по всей видимости, напрямую зависит от физических факторов среды. Изучение спектров изменчивости в распределении зоопланктона показало, что причины неоднородности могут быть гораздо более разнообразными и не столь однозначными. Так, например, в работе (Steele, Henderson, 1992) на примере простой одномерной модели "хищник - жертва" с диффузией и белым шумом, который добавлен к стационарным значениям параметров, показана возможность получения наблюдаемого спектра изменчивости фито- и зоопланктона. Другими словами, было выдвинуто предположение о том, что пятнистость распределения планктона не требует для своего объяснения привлечения сложных гипотез о поведении зоопланктона, а генерируется внутренними нелинейными механизмами, представленными в модели.
С помощью спутникового дистанционного зондирования океана показано, что наблюдаемый спектр флуктуаций концентрации фитопланктона пропорционален ![]() , где k - волновое число. Другими исследованиями установлено, что для квазигеострофических течений энергетический спектр флуктуаций пропорционален
, где k - волновое число. Другими исследованиями установлено, что для квазигеострофических течений энергетический спектр флуктуаций пропорционален ![]() . Аналогичная закономерность установлена и для спектра температурных флуктуаций. На основании этого выдвигается гипотеза о том, что фитопланктон можно рассматривать как пассивную субстанцию, переносимую океанскими течениями, а пятнистость распределения определяется пространственной неоднородностью океанологических полей (Gower et al., 1980). Критика этой гипотезы содержится в работе (Lesieur, Sadourny, 1981). Поводом для этого служит то, что клетки фитопланктона непрерывно репродуцируют, концентрация хлорофилла, следовательно, не является консервативной субстанцией. Поэтому не совсем ясно, как это влияет на форму спектра. Кроме того, теряет смысл использование термина "пассивная (консервативная) субстанция".
. Аналогичная закономерность установлена и для спектра температурных флуктуаций. На основании этого выдвигается гипотеза о том, что фитопланктон можно рассматривать как пассивную субстанцию, переносимую океанскими течениями, а пятнистость распределения определяется пространственной неоднородностью океанологических полей (Gower et al., 1980). Критика этой гипотезы содержится в работе (Lesieur, Sadourny, 1981). Поводом для этого служит то, что клетки фитопланктона непрерывно репродуцируют, концентрация хлорофилла, следовательно, не является консервативной субстанцией. Поэтому не совсем ясно, как это влияет на форму спектра. Кроме того, теряет смысл использование термина "пассивная (консервативная) субстанция".
Еще один подход к анализу изменчивости спектра рассмотрен в работе (Denman, Platt, 1976). Предположим, что турбулентные вихри характеризуются масштабом длины L (волновое число k ![]() ) и соответствующим временным масштабом
) и соответствующим временным масштабом ![]() . За масштаб времени выбрано время, в течение которого кинетическая энергия от вихрей, размером L, переносится к турбулентным вихрям вдвое меньшего размера. Суть предлагаемого метода состоит в том, чтобы сравнить этот временной масштаб
. За масштаб времени выбрано время, в течение которого кинетическая энергия от вихрей, размером L, переносится к турбулентным вихрям вдвое меньшего размера. Суть предлагаемого метода состоит в том, чтобы сравнить этот временной масштаб ![]() с характерным временным масштабом
с характерным временным масштабом ![]() - временем размножения фитопланктона (
- временем размножения фитопланктона (![]() - экспоненциальная скорость роста). В соответствии с амплитудами
- экспоненциальная скорость роста). В соответствии с амплитудами ![]() и
и ![]() можно выделить три характерных режима:
можно выделить три характерных режима:
(1) ![]() . В этом случае неоднородности пространственного распределения фитопланктона будут быстро разрушаться под действием турбулентного перемешивания. Размножение имеет пренебрежимо малый эффект. Ожидаемая форма спектра для хлорофилла не будет иметь значительных отличий от спектра, характерного для консервативной субстанции, такой, например, как температура;
. В этом случае неоднородности пространственного распределения фитопланктона будут быстро разрушаться под действием турбулентного перемешивания. Размножение имеет пренебрежимо малый эффект. Ожидаемая форма спектра для хлорофилла не будет иметь значительных отличий от спектра, характерного для консервативной субстанции, такой, например, как температура;
(2) ![]() . В этом случае размножение фитопланктона играет важную роль и следует ожидать, что при этом режиме на всех масштабах длин будут иметь место существенные отличия в структуре спектров фитопланктона и температуры;
. В этом случае размножение фитопланктона играет важную роль и следует ожидать, что при этом режиме на всех масштабах длин будут иметь место существенные отличия в структуре спектров фитопланктона и температуры;
(3) ![]() . Данная область временных масштабов является переходной между режимами (1) и (2).
. Данная область временных масштабов является переходной между режимами (1) и (2).
Рассмотрим поле турбулентного потока, которое является пространственно изотропным, а турбулентность генерируется только на достаточно больших пространственных масштабах. В этом диапазоне существует область волновых чисел (называемая инерционной областью), в пределах которой флуктуации скорости переносятся без потерь кинетической энергии от низких волновых чисел к более высоким волновым числам до тех пор, пока не будут достигнуты достаточно маленькие масштабы, на которых становится существенным влияние вязкой диссипации (Колмогоров, 1941). Обозначим через ![]() одномерный спектр турбулентной энергии, где символ v показывает, что
одномерный спектр турбулентной энергии, где символ v показывает, что ![]() представляет собой изменчивость флуктуаций скорости. В пределах инерционной области
представляет собой изменчивость флуктуаций скорости. В пределах инерционной области ![]() зависит только от волнового числа k и от скорости переноса кинетической энергии от более низких к более высоким волновым числам. Поскольку для этого каскада выполняется закон сохранения энергии, то есть он является консервативным, то скорость переноса энергии должна быть в точности равна скорости вязкой диссипации
зависит только от волнового числа k и от скорости переноса кинетической энергии от более низких к более высоким волновым числам. Поскольку для этого каскада выполняется закон сохранения энергии, то есть он является консервативным, то скорость переноса энергии должна быть в точности равна скорости вязкой диссипации ![]() на малых масштабах. Анализ размерностей, основанный на этих предположениях, приводит к хорошо известному соотношению (Колмогоров, 1941):
на малых масштабах. Анализ размерностей, основанный на этих предположениях, приводит к хорошо известному соотношению (Колмогоров, 1941):
![]() , (1)
, (1)
где А - безразмерная постоянная. Справедливость соотношения (1) была проверена для районов океана, в которых турбулентное перемешивание характеризуются достаточно большими числами Рейнольдса.
Рассмотрим, далее, скалярную переменную, например такую, как температура ![]() . Нам понадобится один дополнительный параметр для того, чтобы можно было определить спектр ее изменчивости
. Нам понадобится один дополнительный параметр для того, чтобы можно было определить спектр ее изменчивости ![]() , а именно
, а именно ![]() , который представляет собой скорость разрушения спектра температуры при больших волновых числах. Этот параметр по своему физическому смыслу полностью аналогичен
, который представляет собой скорость разрушения спектра температуры при больших волновых числах. Этот параметр по своему физическому смыслу полностью аналогичен ![]() . Анализ размерностей для инерционного диапазона показывает, что изменение
. Анализ размерностей для инерционного диапазона показывает, что изменение ![]() , также как и изменение
, также как и изменение ![]() , будет пропорционально
, будет пропорционально ![]() (Обухов, 1949):
(Обухов, 1949):
![]() , (2)
, (2)
где ![]() - еще одна безразмерная постоянная.
- еще одна безразмерная постоянная.
Предположим, что спектральное распределение ![]() обилия фитопланктона
обилия фитопланктона ![]() в условиях океанской турбулентности будет зависеть не только от параметров
в условиях океанской турбулентности будет зависеть не только от параметров ![]() , но также и от параметра удельной скорости роста фитопланктона
, но также и от параметра удельной скорости роста фитопланктона ![]() . Тот же анализ размерностей дает в этом случае, что
. Тот же анализ размерностей дает в этом случае, что
![]() . (3)
. (3)
Поскольку мы имеем неизвестных больше, чем число уравнений, то F представляет собой неопределенную функцию безразмерной комбинации ![]() . Введем в рассмотрение характеристическое волновое число
. Введем в рассмотрение характеристическое волновое число ![]() , которое можно рассматривать как такой масштаб, на котором рост фитопланктона и диссипация турбулентной кинетической энергии одинаково важны в процессе формирования пространственного распределения фитопланктона. Тогда соотношение (3) можно переписать следующим образом:
, которое можно рассматривать как такой масштаб, на котором рост фитопланктона и диссипация турбулентной кинетической энергии одинаково важны в процессе формирования пространственного распределения фитопланктона. Тогда соотношение (3) можно переписать следующим образом:
![]() . (4)
. (4)
Для волновых чисел ![]() можно ожидать, что
можно ожидать, что ![]() будет незначительно влиять на
будет незначительно влиять на ![]() и поэтому форма спектра изменчивости фитопланктона в этом диапазоне волновых чисел будет аналогична спектру температуры
и поэтому форма спектра изменчивости фитопланктона в этом диапазоне волновых чисел будет аналогична спектру температуры ![]() . Однако, для
. Однако, для ![]() уже скорость диссипации турбулентной кинетической энергии
уже скорость диссипации турбулентной кинетической энергии ![]() не будет влиять на спектр и в этом случае предполагается , что
не будет влиять на спектр и в этом случае предполагается , что ![]() будет иметь следующий, более простой, вид:
будет иметь следующий, более простой, вид:
![]() , (5)
, (5)
где С - безразмерная постоянная.
Пределы изменчивости ![]() можно оценить, исходя из наших знаний порядков величин
можно оценить, исходя из наших знаний порядков величин ![]() и
и ![]() . Для активно растущего фитопланктона скорость роста
. Для активно растущего фитопланктона скорость роста ![]() меняется в пределах от 0,5 до 3,0?10-5 с-1. В пределах верхнего 50-метрового слоя океана диапазон скоростей диссипации турбулентной кинетической энергии составляет 10-7 - 10-9 м2 с-3. Используя эти значения, можно получить, что
меняется в пределах от 0,5 до 3,0?10-5 с-1. В пределах верхнего 50-метрового слоя океана диапазон скоростей диссипации турбулентной кинетической энергии составляет 10-7 - 10-9 м2 с-3. Используя эти значения, можно получить, что ![]() меняется от 0,2 до 20 км. Для средних условий можно ожидать, что
меняется от 0,2 до 20 км. Для средних условий можно ожидать, что ![]() - порядка 1 км.
- порядка 1 км.
По поводу вывода формулы (5) можно сделать несколько критических замечаний. Прежде всего следует обратить внимание на то, что турбулентные движения в пределах верхнего слоя океана не являются ни трехмерными, ни изотропными. Так, наблюдения спектра изменчивости температуры морской поверхности на этих масштабах показывают, что он пропорционален ![]() , а не
, а не ![]() . Если спектр пропорционален
. Если спектр пропорционален ![]() , то это свидетельствует о том, что турбулентный перенос происходит в области фронтальных зон или неоднородностей. Если же он пропорционален
, то это свидетельствует о том, что турбулентный перенос происходит в области фронтальных зон или неоднородностей. Если же он пропорционален ![]() , то мы, по всей видимости, имеем дело с двумерным полем турбулентности.
, то мы, по всей видимости, имеем дело с двумерным полем турбулентности.
При описании, математическом моделировании и исследованиях устойчивости экосистем, в частности озерных, предполагается, что математическая структура уравнений аналогична структуре уравнений химической кинетики: временные зависимости концентраций продуктов определяются локальными значениями концентраций субстратов и продуктов. Однако, такое предположение может приводить к качественному противоречию с экспериментом. Это может иметь место, например, в том случае, если масштабы пространственных неоднородностей компонентов экосистемы меньше или соизмеримы с характерными размерами самой системы. Устойчивость экосистемы в таком случае определяется нелинейным взаимодействием гидродинамического движения с той или иной популяцией и не может быть исследована путем изучения "средних" популяционных уровней. При измерении тех или иных параметров экосистем знание масштабов пространственно-временных неоднородностей, очевидно, необходимо для объективного анализа получаемых в дискретном эксперименте величин.
В работе (Верхозина и др., 1988) путем измерений и анализа пространственной неоднородности в распределении хемоорганотрофных бактерий показано, что такая, нетипичная для обычных озер, ситуация реализуется на Байкале. Низкая скорость размножения типичных биологических видов и достаточная высокая скорость турбулентного перемешивания определяют характерный масштаб "пятнистой" структуры пространственного распределения видов.
Из простых физических соображений ясно, что достаточно мелкие организмы, такие как некоторые виды фитопланктона или, как в данной работе, микроорганизмы должны двигаться в воде как пассивная молекулярная примесь, поскольку скорость их броуновского движения или другого "случайного" блуждания превышает некоторую характерную скорость, например, скорость гравитационного оседания. Простые оценки показывают, что размер таких частиц не должен превышать 10-3 см. Для оценки масштабов пространственных неоднородностей, концентрации биологических объектов при взаимодействии их с турбулентным движением воды, необходимо установить вид спектра неоднородностей, то есть характерных размеров "пятен" и расстояний между "пятнами". В работе (Denman et al., 1977) методами теории химически реагирующих в турбулентном потоке смесей получено выражение для формы спектра флуктуаций концентраций фитопланктона как функции волнового числа k (то есть числа элементов пятнистой структуры в единице длины). Спектр получен как решение гидродинамических уравнений для области волновых чисел, соответствующих инерционной области турбулентного движения, где можно пренебречь эффектами молекулярной диффузии. Этот спектр качественно отличен от ранее полученного в работе (Denman, Platt, 1976) спектра методами анализа размерностей. Отличие состоит в том, что он имеет максимум при волновом числе ![]() :
:
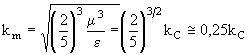 , (6)
, (6)
где, как и раньше, ![]() - скорость экспоненциального роста концентрации фитопланктона,
- скорость экспоненциального роста концентрации фитопланктона, ![]() - скорости вязкой диссипации турбулентной кинетической энергии в единице массы жидкости,
- скорости вязкой диссипации турбулентной кинетической энергии в единице массы жидкости, ![]() - характеристическое волновое число (
- характеристическое волновое число (![]() ). Спектр, полученный в (Denman, Platt, 1976), при
). Спектр, полученный в (Denman, Platt, 1976), при ![]() не имеет максимума: он монотонно возрастает пропорционально
не имеет максимума: он монотонно возрастает пропорционально ![]() . Предполагалось (Denman et al., 1977), что указанное различие возникло из-за того, что при анализе размерностей в области
. Предполагалось (Denman et al., 1977), что указанное различие возникло из-за того, что при анализе размерностей в области ![]() пренебрегали конвективными эффектами (то есть членами, содержащими
пренебрегали конвективными эффектами (то есть членами, содержащими ![]() ) по сравнению с влиянием роста биомассы (то есть членами, содержащими
) по сравнению с влиянием роста биомассы (то есть членами, содержащими ![]() ), поэтому спектр при
), поэтому спектр при ![]() и не зависел от
и не зависел от ![]() .
.
Вид спектра флуктуаций планктона (и любых других неконсервативных частиц) может быть определен без всяких упрощающих предположений на основе метода анализа размерностей развитого турбулентного движения для инерционной области. Для этого достаточно воспользоваться известной формулой для скорости расхождения частиц на расстояние ![]() в инерционной области турбулентной диффузии:
в инерционной области турбулентной диффузии:
![]() . (7)
. (7)
Будем считать задачу двумерной, то есть полагаем, что фитопланктон распределен и перемещается вдоль поверхности воды. Запишем уравнение неразрывности для консервативной примеси:
![]() , (8)
, (8)
где ![]() ,
, ![]() - текущая и начальная концентрации планктона. Тогда получим:
- текущая и начальная концентрации планктона. Тогда получим:
![]() . (9)
. (9)
Если концентрация фитопланктона увеличивается со скоростью ![]() , где
, где ![]() - концентрация фитопланктона, при которой его размножение прекращается ("равновесная" концентрация), то уравнение (9) можно переписать следующим образом:
- концентрация фитопланктона, при которой его размножение прекращается ("равновесная" концентрация), то уравнение (9) можно переписать следующим образом:
![]() . (10)
. (10)
Если перейти к волновым числам, то получим
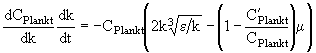 . (11)
. (11)
Из (11) непосредственно следует, что при ![]() зависимость
зависимость ![]() имеет максимум при
имеет максимум при 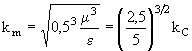 , то есть при практически том же значении, что и (6). При
, то есть при практически том же значении, что и (6). При ![]() максимум вырождается, и зависимость
максимум вырождается, и зависимость ![]() становится монотонной, приближаясь к таковой для консервативной примеси. Интегрируя (11) при начальном волновом числе источника
становится монотонной, приближаясь к таковой для консервативной примеси. Интегрируя (11) при начальном волновом числе источника ![]() , получаем окончательно
, получаем окончательно
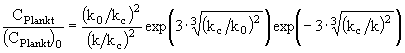 . (12)
. (12)
Таким образом, анализ размерностей позволяет последовательным образом построить зависимость ![]() для неконсервативных частиц и, следовательно, получить спектр их распределения. Следует, однако, заметить, что условие
для неконсервативных частиц и, следовательно, получить спектр их распределения. Следует, однако, заметить, что условие ![]() , использованное при оценке масштаба неоднородностей, в природе может и не соблюдаться.
, использованное при оценке масштаба неоднородностей, в природе может и не соблюдаться.
Оценки неоднородностей распределения хемоорганотрофных бактерий, полученные по формуле (11), хорошо согласуются с наблюдаемыми масштабами пространственных неоднородностей для озера Байкал. Таким образом, проведенное теоретическое и экспериментальное исследование свидетельствуют о наличии существенных пространственных неоднородностей в экосистеме озера. Они связаны с влиянием гидродинамических параметров на структуру экосистемы таким же образом, как в морях и океанах.
В работах (Webster, 1990; Verhagen, 1994; Webster, Hutchinson, 1994) предлагается несколько однотипных по своей структуре моделей, которые описывают формирование пятнистого распределения фитопланктона под влиянием ветровой циркуляции в озерах. Показано, что в достаточно больших озерах горизонтальное распределение фитопланктона определяется ветровыми ситуациями и пространственные структуры могут существовать до нескольких недель. Вертикальная структура меняется гораздо быстрее - в пределах одного дня. Все эти модели являются нестационарными моделями и имеет двухслойную структуру. В верхнем слое течение совпадает с направлением ветра, в нижнем - направление течения противоположно направлению ветра. Отдельно рассматриваются процессы вертикального перемешивания (подъем или опускание вод) вблизи над- и подветренного участков берега. Получены соответствующие формулы для расчета вертикального адвективного транспорта клеток фитопланктона с учетом скорости их плавания. Эта группа моделей также рассматривает фитопланктон как консервативную субстанцию. При таком подходе не учитываются процессы роста, размножения, трофические взаимодействия в планктонном сообществе и условия биогенного питания фитопланктона и светового лимитирования.
В работе (Evans, Taylor, 1980) рассматривается еще один, чисто гидрофизический, процесс формирования пэтчинга планктона - аккумуляция фитопланктона в ячейках Ленгмюра. Ячейки Ленгмюра представляют собой трубки вращающейся воды, их оси выстраиваются примерно вдоль ветра, соседние ячейки вращаются в противоположных направлениях. Аккумуляция фитопланктона будет иметь место вдоль линий конвергенции воды. Это не самый важный механизм аккумуляции, поскольку время существования подобных ячеек достаточно короткое и они имеют небольшие пространственные масштабы (порядка нескольких метров - первых десятков метров), в то время как другие механизмы могут формировать пространственные неоднородности с масштабом в несколько километров. Вертикальные размеры ячеек могут меняться от нескольких сантиметров до сотен метров. Обычно ячейки в поперечнике имеют ширину порядка 10 метров, максимальную поверхностную скорость 1,5 см/с. Такие циркуляционные образования могут иметь место при скоростях ветра порядка 5 м/с. Предлагается очень простая модель аккумуляции фитопланктона в ячейках Ленгмюра на основе модели Стоммела (Stommel, 1949). Показано, что данная модель перестает работать, если собственная скорость перемещения планктонных организмов не является постоянной величиной.
Концентрация фитопланктона в воде зависит от большого числа факторов: концентрации биогенных элементов, температуры воды, освещенности, прозрачности воды, сезона года и т.д. Кроме того, скопления планктонных организмов переносятся движущимися водными массами. В связи с малой подвижности клеток фитопланктона их часто можно рассматривать как примесь, пассивно переносимую жидкостью (Bennett, Denman, 1985). В свою очередь, такой перенос примеси можно разделить на две составляющие: перенос, обусловленный полем осредненных скоростей (адвекция), и перенос, вызванный наличием в поле скорости случайных, хаотических составляющих (турбулентная диффузия). Адвекция переносит скопления примеси как целое, не влияя существенным образом на внутреннюю структуру скоплений. В противоположность этому, диффузия активно воздействует на структуру скоплений, стремясь выровнять существующие в поле концентрации примеси градиенты. Картина усложняется в случае неконсервативной примеси, когда ее количество может уменьшаться или, наоборот, увеличиваться со временем.
В работе (Озмидов, 1998) показано, что пятна фитопланктона могут существовать и эволюционировать в условиях однородности, стационарности и горизонтальной изотропности океанологических полей, а также стационарности биологических параметров. Запишем уравнение, описывающее изменение численности фитопланктона:
![]() , (13)
, (13)
где ![]() - скорость фотосинтеза на единицу количества организмов, R - скорость дыхания, G - скорость выедания фитопланктона зоопланктоном. При рассмотрении процесса диффузии скоплений фитопланктона в течение небольших отрезков времени многие факторы, ограничивающие развитие клеток, не существенны, в связи с чем величину
- скорость фотосинтеза на единицу количества организмов, R - скорость дыхания, G - скорость выедания фитопланктона зоопланктоном. При рассмотрении процесса диффузии скоплений фитопланктона в течение небольших отрезков времени многие факторы, ограничивающие развитие клеток, не существенны, в связи с чем величину ![]() можно считать постоянной величиной. В этом случае в системе координат, движущейся вместе с осредненным течением, можно записать уравнение баланса количества фитопланктона (начало координат расположено на поверхности океана, вертикальная ось направлена вниз):
можно считать постоянной величиной. В этом случае в системе координат, движущейся вместе с осредненным течением, можно записать уравнение баланса количества фитопланктона (начало координат расположено на поверхности океана, вертикальная ось направлена вниз):
![]() , (14)
, (14)
где ![]() - концентрация фитопланктона,
- концентрация фитопланктона, ![]() ,
, ![]() ,
, ![]() - коэффициенты турбулентного обмена вдоль соответствующих осей координат, t - время.
- коэффициенты турбулентного обмена вдоль соответствующих осей координат, t - время.
Специфика турбулентности, обеспечивающей диффузию примеси, характеризуется величиной и функциональным видом коэффициентов турбулентного обмена. В данной работе рассматриваются четыре типичных режима турбулентности:
1) Этот режим следует ожидать в динамически спокойных районах океана при тихой погоде с прогревом верхнего водного слоя Солнцем и с образованием высоко расположенного суточного скачка плотности воды. Вертикальный обмен в этих условиях затруднен. Горизонтальный обмен, предполагаемый однородным и изотропным, обусловливается мелкомасштабной турбулентностью, что описывается постоянными коэффициентами обмена ![]() .
.
2) Этот режим типичен для динамически спокойных районов океана с относительно глубоко расположенным слоем скачка плотности при свежей погоде. Турбулентность в верхнем перемешанном слое трехмерна, мелкомасштабна и горизонтально изотропна, то есть ![]() ,
, ![]() .
.
3) Режим характерен для динамически активных районов океана (фронтальные зоны, конвергенции и дивергенции течений, зоны вихреобразования и т. п.) с четко выраженным слоем скачка плотности. Турбулентность горизонтально однородна, изотропна и многомасштабна, то есть коэффициенты ![]() зависят от масштаба явления и могут быть выражены через радиус-вектор r по формуле закона "степени 4/3":
зависят от масштаба явления и могут быть выражены через радиус-вектор r по формуле закона "степени 4/3":
![]() , (15)
, (15)
где ![]() - скорость диссипации турбулентной энергии,
- скорость диссипации турбулентной энергии, ![]() - постоянная. Для верхнего слоя океана
- постоянная. Для верхнего слоя океана ![]() = 10-2 см2/с3,
= 10-2 см2/с3, ![]() = 0,1.
= 0,1.
4) Этот режим может существовать в очень специфических районах океана в динамически активных зонах с развитыми вертикальными движениями вод, например, в районах глубокой зимней конвекции. Вертикальный градиент плотности отсутствует, трехмерная изотропная турбулентность многомасштабна, коэффициент обмена может быть задан формулой ![]() , где R - трехмерный радиус-вектор.
, где R - трехмерный радиус-вектор.
Применительно к внутренним водоемам нас больше всего будет интересовать первый режим турбулентности, который мы и рассмотрим более подробно. При сделанных выше предположениях о виде коэффициентах турбулентного обмена уравнение переноса примеси (14) перепишется в полярных координатах следующим образом:
![]() . (16)
. (16)
Фундаментальное решение этого классического уравнения Фика для двумерной диффузии известно и имеет следующий вид:
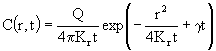 . (17)
. (17)
Здесь Q - мгновенный точечный источник примеси, помещенный в начальный момент времени в начало координат. Проанализируем поведение пятна во времени в зависимости от значений параметров, входящих в решение. Как видно из решения, концентрация примеси в пятне зависит от времени диффузии t и расстояния точки наблюдения от центра пятна r. Концентрация примеси в центре пятна (r = 0) в случае консервативной примеси (![]() = 0) меняется со временем обратно пропорционально времени диффузии. Для неконсервативной примеси (
= 0) меняется со временем обратно пропорционально времени диффузии. Для неконсервативной примеси (![]() > 0) эта зависимость более сложна и определяется соотношением множителей
> 0) эта зависимость более сложна и определяется соотношением множителей ![]() и
и ![]() . Для исследования зависимости размера пятна от времени диффузии прежде всего определим, что мы будем понимать под размером пятна. Радиус пятна определяется по расстоянию от центра пятна до некоторой изолированной концентрации
. Для исследования зависимости размера пятна от времени диффузии прежде всего определим, что мы будем понимать под размером пятна. Радиус пятна определяется по расстоянию от центра пятна до некоторой изолированной концентрации ![]() , вне которой концентрацией примеси пренебрегается. При таком предположении определим зависимость радиуса пятна от времени диффузии из формулы (17). Прологарифмируем это соотношение и проведем некоторые алгебраические преобразования. Получим:
, вне которой концентрацией примеси пренебрегается. При таком предположении определим зависимость радиуса пятна от времени диффузии из формулы (17). Прологарифмируем это соотношение и проведем некоторые алгебраические преобразования. Получим:
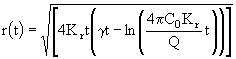 . (18)
. (18)
Вначале рассмотрим случай консервативной примеси, когда (![]() = 0). Зависимость радиуса пятна от времени t удобно рассмотреть в безразмерных координатах. Для этого введем безразмерные переменные по формулам:
= 0). Зависимость радиуса пятна от времени t удобно рассмотреть в безразмерных координатах. Для этого введем безразмерные переменные по формулам:
![]() , (19)
, (19)
![]() . (20)
. (20)
Старые переменные будут при этом выражаться через новые по формулам:
![]() , (21)
, (21)
![]() . (22)
. (22)
В новых безразмерных переменных зависимость радиуса пятна консервативной примеси от времени диффузии выражается компактной формулой:
![]() . (23)
. (23)
Отсюда следует, что в рассматриваемом случае радиус пятна вначале быстро растет, достигает максимальной величины при значении ![]() , равном приблизительно 0,4, затем убывает и при
, равном приблизительно 0,4, затем убывает и при ![]() пропадает, то есть концентрация планктона становится меньше значения
пропадает, то есть концентрация планктона становится меньше значения ![]() во всем рассматриваемом пространстве. Длительность существования пятна зависит, естественно, от значения определяющих параметров Q,
во всем рассматриваемом пространстве. Длительность существования пятна зависит, естественно, от значения определяющих параметров Q, ![]() и
и ![]() . Более сложной оказывается зависимость радиуса пятна от времени для неконсервативной примеси. В этом случае становится важным соотношение членов
. Более сложной оказывается зависимость радиуса пятна от времени для неконсервативной примеси. В этом случае становится важным соотношение членов ![]() и
и ![]() в выражении (18). Так как величины Q,
в выражении (18). Так как величины Q, ![]() ,
, ![]() , t и
, t и ![]() существенно положительны, то при
существенно положительны, то при ![]() оба рассматриваемых члена дают положительный вклад в подкоренное выражение формулы (18). При
оба рассматриваемых члена дают положительный вклад в подкоренное выражение формулы (18). При ![]() логарифмический член дает отрицательный вклад, однако положительное значение подкоренной функции может сохраняться и далее в зависимости от взаимного расположения графиков линейной и логарифмической функции. Анализ показывает, что в случае неконсервативной примеси и при двумерной мелкомасштабной турбулентности теоретически могут существовать четыре режима "жизни" диффундирующих пятен фитопланктона: непрерывный с длительным существованием пятна, но с переменной скоростью его роста (или уменьшения), пульсирующий, при котором первоначальное пятно исчезает в некоторый момент времени, затем вновь возникает и начинает расти, третий - перемежающийся, когда пятно исчезает, отсутствует некоторое время, после чего появляется снова, и, наконец, четвертый, когда возникшее пятно достаточно быстро исчезает и не появляется вновь длительное время. Реальный режим, как показывают приведенные в данной работе расчеты, зависит от соотношения параметров и, самое главное, от масштаба времени, в пределах которого справедливо допущение о постоянстве коэффициента размножения
логарифмический член дает отрицательный вклад, однако положительное значение подкоренной функции может сохраняться и далее в зависимости от взаимного расположения графиков линейной и логарифмической функции. Анализ показывает, что в случае неконсервативной примеси и при двумерной мелкомасштабной турбулентности теоретически могут существовать четыре режима "жизни" диффундирующих пятен фитопланктона: непрерывный с длительным существованием пятна, но с переменной скоростью его роста (или уменьшения), пульсирующий, при котором первоначальное пятно исчезает в некоторый момент времени, затем вновь возникает и начинает расти, третий - перемежающийся, когда пятно исчезает, отсутствует некоторое время, после чего появляется снова, и, наконец, четвертый, когда возникшее пятно достаточно быстро исчезает и не появляется вновь длительное время. Реальный режим, как показывают приведенные в данной работе расчеты, зависит от соотношения параметров и, самое главное, от масштаба времени, в пределах которого справедливо допущение о постоянстве коэффициента размножения ![]() . Если промежуток времени, на котором анализируется модель, превышает этот масштаб, то поведение пятна не может быть адекватно описано в рамках рассматриваемой модели. Безразмерное выражение для радиуса диффундирующего пятна при
. Если промежуток времени, на котором анализируется модель, превышает этот масштаб, то поведение пятна не может быть адекватно описано в рамках рассматриваемой модели. Безразмерное выражение для радиуса диффундирующего пятна при ![]() будет иметь вид:
будет иметь вид:
![]() . (24)
. (24)
В случае второго режима уравнение переноса примеси имеет следующий вид:
 . (25)
. (25)
Его решение также известно как решение Фика для трехмерной диффузии в полупространстве ![]() :
:
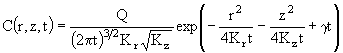 . (26)
. (26)
Таким образом, в отличие от первого режима, концентрация примеси в центре пятна уменьшается со временем обратно пропорционально времени диффузии t в степени 3/2. Выражение для размера пятна фитопланктона в трехмерном случае может быть получено из данного решения (26) аналогично использованному ранее приему для двумерной задачи. Логарифмируя (26) и решая полученное уравнение относительно r, будем иметь для поверхности океана:
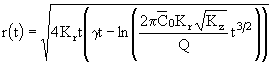 , (27)
, (27)
где ![]() - пороговая объемная концентрация фитопланктона. К безразмерному виду соотношение (27) приводится с помощью формул:
- пороговая объемная концентрация фитопланктона. К безразмерному виду соотношение (27) приводится с помощью формул:
![]() , (28)
, (28)
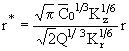 . (29)
. (29)
Обратные преобразования координат задаются выражениями:
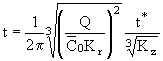 , (30)
, (30)
 . (31)
. (31)
В безразмерных координатах выражение (27) приобретает вид:
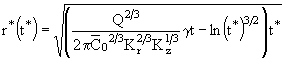 . (32)
. (32)
В случае консервативной примеси формула (32) упрощается и приводится к компактному виду:
![]() . (33)
. (33)
С помощью численных оценок было показано, что в случае быстро размножающегося планктона поведения пятна в трехмерном случае сильно отличается от соответствующей зависимости для двумерного случая. Усилившаяся в трехмерном случае диффузия переводит режим существования пятна из непрерывного в перемежающийся. Что касается слабо размножающегося планктона, то поведение его скоплений практически неотличимо от поведения консервативной примеси.
Режим турбулентности самым существенным образом влияет на поведение пятен примеси в океане. Было показано, что увеличение интенсивности турбулентности (переход от режима 1 к последующим режимам) в целом снижает значения радиусов пятен и времени их существования. В то же время пятна быстро размножающегося фитопланктона становятся более схожими с пятнами консервативной примеси (диффузия превалирует над эффектами размножения организмов). Активного развития пятен фитопланктона следует ожидать при первом режиме турбулентности, то есть при тихой погоде в прогреваемом Солнцем верхнем слое океана (естественно, при достаточном количестве биогенных элементов и при наличии других факторов, необходимых для развития организмов). При втором режиме турбулентности пятна фитопланктона могут образовываться эпизодически и существовать сравнительно недолго. Районы с третьим режимом турбулентности не благоприятны для развития пятен фитопланктона, так как они будут быстро разрушаться горизонтальной турбулентностью. Развитие пятен фитопланктона в зонах с четвертым режимом турбулентности вообще маловероятно. Принятая в моделях идеализированная форма пятен в виде окружностей в природных условиях, естественно, нарушается, и пятна примеси имеют более сложную форму. В связи с этим под радиусами пятен в моделях следует понимать радиусы кругов, эквивалентные по площади пятнам диффундирующей примеси.
Учет пространственной неоднородности в математических моделях водных экосистем позволяет изучать их качественные свойства. Диффузионные модели удачно применяются для описания пространственно неоднородных структур в распределении планктонных сообществ (Levin, 1979). Так, в работе Стила (Steele, 1974) предложено исследование устойчивости планктонной системы, состоящей из фитопланктона и растительноядного зоопланктона с учетом горизонтальной неоднородности среды. Анализ модели проведен как в линейном (методом малых возмущений), так и в нелинейном приближении по отношению к учету членов, описывающих взаимодействие фито- и зоопланктона. Оказалось, что если система будет неустойчивой при отсутствии турбулентной диффузии, то в ее присутствии система приобретает устойчивость при малых горизонтальных масштабах процесса (для открытой части моря это составляет 10 - 100 километров). Одновременно с этим была обнаружена корреляция между пространственными колебаниями численностей популяций разных трофических уровней.
Возможность появления пятен фитопланктона в горизонтальном направлении изучено с помощью другой, более детализированной, модели морской экосистемы (Suzuki, Fukuoka, 1979). Модель представляет собой систему трех дифференциальных уравнений, описывающих динамику питательных веществ, фито- и зоопланктона. В предположении, что биогенные вещества и фитопланктон имеют одинаковые коэффициенты диффузии, авторы показали, что диффузионная неустойчивость возникает в том случае, когда зоопланктон имеет коэффициент диффузии в 100 раз больший, чем фитопланктон, что вряд ли возможно в реальных экосистемах. Совместное действие таких двух факторов, как диффузия и трофические взаимоотношения между фито- и зоопланктоном, способно привести к неоднородности их пространственного распределения. Общая теория возникновения диффузионной неустойчивости и методы исследования систем с диффузией приведены в работах (Романовский и др., 1984.; Свирежев, Логофет, 1978; Марри, 1983; Свирежев, 1987), в которых теоретические результаты иллюстрируются конкретными биологическими примерами. Диффузионная неустойчивость в математических моделях вертикально распределенных водных экосистем исследована в работе (Тонких, 1985).
В классической работе (Kierstead, Slobodkin, 1953), как уже отмечалось выше, рассматривались не только физические, но также и биологические процессы, происходящие в пределах фитопланктонного пятна. Было показано, что неоднородность в распределении планктона возникает в результате нелинейного взаимодействия квазидетерминированных биологических процессов роста, смертности, трофических взаимодействий (по типу хищник-жертва) с мелкомасштабной турбулентностью в океане. Получено соотношение между характерным масштабом длины пятна, коэффициентом турбулентной диффузии и скоростью роста фитопланктона, которая требуется для того, чтобы сбалансировать потери в результате диффузионного рассеяния:
![]() , (34)
, (34)
где ![]() - размер пятна,
- размер пятна, ![]() - коэффициент турбулентной диффузии и
- коэффициент турбулентной диффузии и ![]() - скорость роста фитопланктона. Например, если
- скорость роста фитопланктона. Например, если ![]() = 10-5 сек-1,
= 10-5 сек-1, ![]() = 106 см2 сек-1, то
= 106 см2 сек-1, то ![]() = 10 км. Пятно меньшего размера не сможет сохраниться и будет размываться под действием турбулентной диффузии. Таким образом,
= 10 км. Пятно меньшего размера не сможет сохраниться и будет размываться под действием турбулентной диффузии. Таким образом, ![]() является "критическим масштабом длины". В более поздних работах рассматривалось влияние выедания фитопланктона нехищным зоопланктоном на критический размер пятна. Предполагая, что выедание происходит в соответствии с функцией Ивлева, была получена уточненная формула для оценки критического масштаба длины
является "критическим масштабом длины". В более поздних работах рассматривалось влияние выедания фитопланктона нехищным зоопланктоном на критический размер пятна. Предполагая, что выедание происходит в соответствии с функцией Ивлева, была получена уточненная формула для оценки критического масштаба длины ![]() :
:
![]() , (35)
, (35)
где R - максимальный рацион нехищного зоопланктона (концентрация сек-1), ![]() - константа Ивлева (конц.-1). Было также показано, что для получения реалистичных решений необходимо рассматривать фактор пищевого лимитирования. Во всех этих ранних и достаточно простых моделях размеры пятна неограниченно увеличивались с течением времени.
- константа Ивлева (конц.-1). Было также показано, что для получения реалистичных решений необходимо рассматривать фактор пищевого лимитирования. Во всех этих ранних и достаточно простых моделях размеры пятна неограниченно увеличивались с течением времени.
В работе (Wroblewski, O'Brien, 1976) развивается концепция критического масштаба длины фитопланктонного пятна и исследуется влияние на размер пятна совместного воздействия турбулентной диффузии, истощения питательных веществ, периодичности световых условий, непрерывного или только ночного выедания фитопланктона зоопланктоном. Физические и биологические процессы, которые определяют концентрацию фитопланктона в произвольной точке океана, можно выразить в форме следующего уравнения:
![]() , (36)
, (36)
где ![]() - концентрация фитопланктона,
- концентрация фитопланктона, ![]() ,
, ![]() ,
, ![]() - коэффициенты турбулентного обмена вдоль соответствующих осей координат, t - время,
- коэффициенты турбулентного обмена вдоль соответствующих осей координат, t - время, ![]() - максимальная удельная скорость роста фитопланктона, S - функция, описывающая процесс роста и учитывающая влияние внешних факторов, QP - слагаемое, которое описывает общие потери фитопланктона в результате смертности, выедания, дыхания, внеклеточных выделений. В данной работе рассматривается только один лимитирующий биогенный элемент N, а функция S выбрана в форме Михаэлиса-Ментен. Таким образом, слагаемое
- максимальная удельная скорость роста фитопланктона, S - функция, описывающая процесс роста и учитывающая влияние внешних факторов, QP - слагаемое, которое описывает общие потери фитопланктона в результате смертности, выедания, дыхания, внеклеточных выделений. В данной работе рассматривается только один лимитирующий биогенный элемент N, а функция S выбрана в форме Михаэлиса-Ментен. Таким образом, слагаемое ![]() принимает следующий вид:
принимает следующий вид: ![]() , где
, где ![]() - константа полунасыщения по азоту. Биомасса фитопланктона и нехищного зоопланктона выражается в единицах лимитирующего биогенного элемента, в данном случае азота. Потери фитопланктона разделяются на два слагаемые: потери, связанные с выеданием зоопланктоном - g(P,Z), и потери, обусловленные внеклеточным выделением и автолизом стареющих клеток - BP. Предполагается, что процесс выедания описывается модифицированной функцией Ивлева:
- константа полунасыщения по азоту. Биомасса фитопланктона и нехищного зоопланктона выражается в единицах лимитирующего биогенного элемента, в данном случае азота. Потери фитопланктона разделяются на два слагаемые: потери, связанные с выеданием зоопланктоном - g(P,Z), и потери, обусловленные внеклеточным выделением и автолизом стареющих клеток - BP. Предполагается, что процесс выедания описывается модифицированной функцией Ивлева:
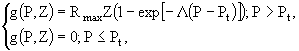 (37)
(37)
где ![]() - максимальная скорость выедания (сек-1),
- максимальная скорость выедания (сек-1), ![]() - постоянная Ивлева,
- постоянная Ивлева, ![]() - пороговая концентрация фитопланктона, ниже которой выедание прекращается. Фактически
- пороговая концентрация фитопланктона, ниже которой выедание прекращается. Фактически ![]() , если только
, если только ![]() .
.
Если для простоты мы будем рассматривать только одно горизонтальное направление Х, постоянное значение коэффициента горизонтальной диффузии ![]() , не будем учитывать влияние адвекции, то получим:
, не будем учитывать влияние адвекции, то получим:
![]() , (38)
, (38)
откуда
![]() . (39)
. (39)
Аналогично, мы можем получить одномерное уравнение для описания временной динамики нехищного зоопланктона Z:
![]() , (40)
, (40)
где увеличение биомассы зоопланктона зависит от функции g(P,Z) и слагаемого, описывающего процесс экскреции - ![]() . Заметим, что коэффициент турбулентной диффузии считается одинаковым как для фитопланктона, так и для зоопланктона, хотя на самом деле для зоопланктона он может быть много меньше или даже может равняться нулю, поскольку взрослые организмы могут сами перемещаться и сохранять плотность пятна.
. Заметим, что коэффициент турбулентной диффузии считается одинаковым как для фитопланктона, так и для зоопланктона, хотя на самом деле для зоопланктона он может быть много меньше или даже может равняться нулю, поскольку взрослые организмы могут сами перемещаться и сохранять плотность пятна.
Уравнение, описывающее процесс диффузии, поглощения и рециклинга лимитирующего биогенного элемента имеет вид:
![]() . (41)
. (41)
Заметим, что если пренебречь воздействием диффузии, то ![]() , то есть выполняется закон сохранения для лимитирующего биогенного элемента.
, то есть выполняется закон сохранения для лимитирующего биогенного элемента.
Формулировка процессов, рассматриваемых в данной модели, в значительной степени упрощена. Например, нереалистично выглядит предположение о немедленной регенерации органических веществ, выделяемых в процессе экскреции, поскольку зоопланктон формирует фекальные пеллеты, которые в результате процесса осаждения могут покидать эвфотическую зону океана. Не рассматриваются также такие важные процессы как формирование, оседание и разложение фекальных пеллет, оседание клеток фитопланктона, вертикальное перемешивание, поглощение света с глубиной.
Перейдем от исходной, размерной, системы уравнений к системе уравнений, записанной в безразмерном виде. Для введения характерного масштаба времени будем использовать максимальную удельную скорость роста ![]() (сек-1). Тогда
(сек-1). Тогда ![]() , где
, где ![]() - безразмерное время. Концентрации P, Z, N выразим через общее количество лимитирующего биогенного элемента в системе
- безразмерное время. Концентрации P, Z, N выразим через общее количество лимитирующего биогенного элемента в системе ![]() :
: ![]() ,
, ![]() ,
, ![]() . Введем также следующие обозначения:
. Введем также следующие обозначения: ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() .
. ![]() - безразмерные концентрации фитопланктона, зоопланктона, биогенов и безразмерная длина соответственно. Тогда уравнения (39) - (41), записанные в безразмерной форме, примут следующий вид:
- безразмерные концентрации фитопланктона, зоопланктона, биогенов и безразмерная длина соответственно. Тогда уравнения (39) - (41), записанные в безразмерной форме, примут следующий вид:
![]() , (42)
, (42)
![]() , (43)
, (43)
![]() . (44)
. (44)
В дальнейшем знак штриха будем опускать.
Для ответа на вопрос о реакции пятна фитопланктона на совместное воздействие турбулентной диффузии, истощения питательных веществ, выедания зоопланктона было проведено несколько серий численных модельных экспериментов на основе системы (42) - (44). При этом были исследованы различные варианты начальных условий, трофических взаимодействий между фито- и зоопланктоном и влияние условий освещенности.
Вариант А:
1) в качестве начальных условий задавалось пятнистое распределение питательных веществ на поверхности океана;
2) предполагалось постоянное выедание фитопланктона зоопланктоном;
3) учитывается влияние турбулентной диффузии.
В этом случае в начальный момент времени размер пятна и концентрация фитопланктона в нем растет (так же как и размер пятна зоопланктона, биогенного элемента), в то время как концентрация лимитирующего питательного элемента убывает. Примерно через восемь суток модельного времени биогенный элемент практически полностью истощается, а постоянно питающийся зоопланктон увеличивается по биомассе за счет фитопланктона. Все три компонента системы постоянно рассеиваются в результате действия турбулентной диффузии. По истечению шестнадцати суток модельного времени остается небольшое пятно фитопланктона (оно не может расти из-за недостатка питательных веществ и усиливающегося выедания и рассеивания), а доминирует пятно зоопланктона. Присутствует незначительное количество биогенного элемента, обусловленное процессом экскреции.
Вариант В:
1) в качестве начальных условий задавалось равномерное распределение питательных веществ без лимитирования;
2) предполагалось постоянное выедание фитопланктона зоопланктоном;
3) учитывается влияние турбулентной диффузии.
В этом случае в начальный момент времени наблюдается активный рост биомассы фитопланктона, что приводит к постепенному локальному истощению биогенного элемента. Размер фитопланктонного пятна увеличивается за счет влияния турбулентной диффузии. Вслед за этим начинается рост пятна зоопланктона и биомассы в нем. Примерно к восьмым суткам модельного времени в результате выедания фитопланктона его пятно расщепляется на две части (две волны), каждая из которых движется в противоположных направлениях в сторону наибольших концентраций биогенного элемента. В центральной части пятна наблюдается практически полное истощение биогенов. Небольшое их количество, как и в варианте А, обусловлено, в основном, процессом экскреции. Пятно зоопланктона также расширяется в результате действия турбулентной диффузии и движения в сторону наибольших концентраций фитопланктона. Таким образом, создается зона резкого градиента питательных веществ на границе пятна фитопланктона, который постепенно уменьшается к центру пятна.
Вариант С:
1) в качестве начальных условий задавалось равномерное распределение питательных веществ без лимитирования;
2) предполагалось постоянное выедание фитопланктона зоопланктоном;
3) введена вертикальная миграция зоопланктона;
4) учитывается влияние турбулентной диффузии.
Для моделирования процесса вертикальной миграции общая биомасса Z зоопланктона разделена на две составляющие. Одна из них, ![]() , постоянно располагается в поверхностных слоях воды и может выедать фитопланктон, другая же часть,
, постоянно располагается в поверхностных слоях воды и может выедать фитопланктон, другая же часть, ![]() , может вертикально мигрировать в более глубокие слои воды, где она не может выедать фитопланктон. Миграция во времени происходит в соответствии с биологическими особенностями данного вида. Уравнение (43), описывающее изменение биомассы зоопланктона, распадается на следующие два уравнения:
, может вертикально мигрировать в более глубокие слои воды, где она не может выедать фитопланктон. Миграция во времени происходит в соответствии с биологическими особенностями данного вида. Уравнение (43), описывающее изменение биомассы зоопланктона, распадается на следующие два уравнения:
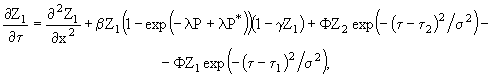 (45)
(45)
![]() , (46)
, (46)
где ![]() - моменты времени в пределах безразмерного времени суток
- моменты времени в пределах безразмерного времени суток ![]() , с которых начинается период подъема или опускания зоопланктона,
, с которых начинается период подъема или опускания зоопланктона, ![]() - доля от безразмерных суток, в пределах которых имеет место вертикальная миграция. Этот период центрируется относительно восхода и захода солнца. Параметр Ф определяет, какая доля от общей биомассы зоопланктона подвержена миграции. Можно предположить, что уменьшение пресса зоопланктона, связанного с миграцией, вызовет рост биомассы фитопланктона. Тем не менее, как показали результаты моделирования, этого в данном случае не происходит. Фактическая стационарная концентрация фитопланктона оказывается ниже по сравнению с вариантом, когда никакой миграции не было. Это происходит из-за устранения части зоопланктона
- доля от безразмерных суток, в пределах которых имеет место вертикальная миграция. Этот период центрируется относительно восхода и захода солнца. Параметр Ф определяет, какая доля от общей биомассы зоопланктона подвержена миграции. Можно предположить, что уменьшение пресса зоопланктона, связанного с миграцией, вызовет рост биомассы фитопланктона. Тем не менее, как показали результаты моделирования, этого в данном случае не происходит. Фактическая стационарная концентрация фитопланктона оказывается ниже по сравнению с вариантом, когда никакой миграции не было. Это происходит из-за устранения части зоопланктона ![]() из биологической динамики в поверхностном слое. Зоопланктон, вертикально мигрирующий в более глубокие слои воды, больше не возобновляет запас питательных веществ в поверхностном слое за счет рециклинга, что приводит к постепенному истощению биогенных элементов. В то же время общая продукция зоопланктона (
из биологической динамики в поверхностном слое. Зоопланктон, вертикально мигрирующий в более глубокие слои воды, больше не возобновляет запас питательных веществ в поверхностном слое за счет рециклинга, что приводит к постепенному истощению биогенных элементов. В то же время общая продукция зоопланктона (![]() +
+![]() ) растет.
) растет.
Вариант D:
1) в качестве начальных условий задавалось равномерное распределение питательных веществ без лимитирования;
2) предполагалось ночное выедание фитопланктона зоопланктоном;
3) введено световое лимитирование скорости роста фитопланктона;
4) учитывается влияние турбулентной диффузии.
В этом случае имели место колебания концентраций фитопланктона и биогенных элементов в пределах пятна.
В классической работе по химическим основам биоморфогенеза Тьюрингом (Turing, 1952) было показано, что диффузия может играть важную роль в формировании пространственных структур. При этом коэффициенты диффузии нелинейно взаимодействующих компонентов должны значительно различаться между собой. Тем не менее, во многих случаях, например в жидких растворах, указанные коэффициенты почти равны. Возникает естественный вопрос: могут ли сформироваться пространственные структуры при таких условиях. Экспериментально проверить идею Тьюринга было трудно даже в случае химических реакций. Это было сделано только в 1990 году. По-видимому, Сигель и Джексон (Segel, Jeckson, 1972) были первыми, кто применил результаты Тьюринга к проблемам популяционной динамики. Ими была показана возможность появления диссипативной неустойчивости при взаимодействии по типу хищник-жертва между фитопланктоном и веслоногими ракообразными с более высокой подвижностью последних. Левин и Сигель (Levin, Segel, 1976) рассматривали этот сценарий образования пространственных структур, как возможный механизм формирования пятнистости планктона. В работе (Rovinsky, Menzinger, 1992) показано, что химическая неустойчивость пространственно однородного распределения может появляться, если реагенты перемещаются с разными скоростями, то есть имеет место дифференциация потоков в разных частях системы. В таком случае в первоначально однородной системе могут формироваться бегущие волны. Аналогичное явление может иметь место и в случае системы взаимодействующих видов, перемещающихся с разными скоростями (Malchow, 2000). Этот механизм формирования пространственных структур является более общим и подходящим по сравнению с механизмом Тьюринга, который накладывает слишком жесткие ограничения на соотношение между коэффициентами диффузии.
В работе (Malchow, 2000) с помощью минимальной модели взаимодействия фито- и зоопланктона демонстрируется диффузионный и адвективный сценарии формирования пространственных структур из первоначально однородного состояния в одно- и двумерном случаях. Было показано, что неустойчивость в первоначально однородном распределении может появляться, если виды движутся с разными скоростями. При этом следует принимать во внимание векторный характер скорости, поскольку виды могут перемещаться, например, с одинаковой по модулю скоростью, но в разных направлениях или же с разными скоростями в одном направлении. Однородное распределение будет оставаться устойчивым для специального случая, когда виды движутся с разными по модулю скоростями в одном направлении и направление вектора волнового возмущения является перпендикулярным к направлению скоростей.
Основная нерешенная проблема в рамках подхода Тьюринга состоит в том, что четкая идентификация активаторов и ингибиторов, которые могли бы принимать участие в формировании самых разных по своей природе структур (физико-химических, биологических или социальных), в большинстве случаев до сих пор отсутствует и, по всей видимости, едва ли может быть реализована. Сегодня получают развитие более реалистические подходы к теории образования структур. Эти подходы рассматривают сложную пространственно-временную динамику открытых пространственно ограниченных систем как результат взаимодействия внутренней динамики таких систем и внешних воздействий, обусловленных влиянием окружающей среды. В экологии такое взаимодействие может включать как физико-химические, так и биологические факторы.
Гетерогенность пространственного распределения различных видов не во всех случаях может сводится к гетерогенности окружающей среды: имеются "физический" и "биологический" масштабы. В работах (Медвинский и др., 2002; Medvinsky et al., 2002) на примере минимальных концептуальных моделей показано, что формирование пространственных структур в распределении планктона, сходных с теми, что наблюдаются в природе (то есть иррегулярных структур с конечным временем жизни), может являться результатом взаимодействия между турбулентными перемешиванием и взаимодействиями между фитопланктоном и зоопланктоном. Формирование планктонной структуры характеризуется наличием присущего ей пространственного масштаба. Его величина (которая по оценкам составляет около 1 км) хорошо согласуется с данными полевых наблюдений. Минимальная модель, достаточная для описания процесса формирования иррегулярного пространственно неоднородного распределения планктона, представляет собой двухкомпонентную систему типа "хищник-жертва" ("зоопланктон" - "фитопланктон") с параметрами, не зависящими от координат, и без каких бы то ни было ограничений на коэффициенты диффузии составляющих систему компонент. Показано, что динамика системы, соответствующая формированию пространственных структур, может быть квалифицирована как пространственно-временной хаос. Появлению (в ходе необычного сценария развития) иррегулярных пространственных распределений может предшествовать формирование хорошо различимых спиральных структур. Показано также, что еще один механизм образования иррегулярных пространственных структур в планктонном сообществе может быть связан с влиянием косяка питающейся планктоном рыбы. Взаимодействие между подвижными рыбными косяками и планктонной системой хотя и модифицирует свойства пространственных структур, но не изменяет основные особенности динамики системы: формирование спиральных волн и хаос в динамике планктона. Показано, что движение рыбного косяка обладает фрактальными свойствами. Во многих случаях динамика водного сообщества подвержена влиянию относительно устойчивых мезомасштабных неоднородностей полей экологически значимых факторов, например, температуры воды, солености, концентрации биогенов. В этом случае характеристический размер этих неоднородностей задает еще одну, внешнюю шкалу системы.
Полученные результаты могут указывать на первостепенную роль хаотических режимов в формировании пространственно-временной организации водных экосистем (Medvinsky et al., 2002). Имеется все возрастающее количество наблюдений в пользу того, что системы с хаотической динамикой могут даже лучше адаптироваться к изменениям окружающих условий, чем системы, динамика которых характеризуется наличием устойчивой точки равновесия.
В работе (Malchow et al., 2002) с помощью минимальной модели пищевой цепи (Scheffer, 1991) (питательные вещества (биоген) R фитопланктон R зоопланктон R рыбы) рассматриваются воздействие подвижности рыбных стай и шума в показателе смертности зоопланктона на динамику планктонного сообщества. Отличительной особенностью предлагаемой модели является то, что рост планктона, его трофодинамические взаимоотношения и перемещение моделируются с помощью системы уравнений типа реакция-диффузия, в то время как для описания движение рыбного косяка используются идеи индивидуально-ориентированного моделирования. Движение стаи рыб в данном случае представляет собой дискретный, основанный на определенных правилах поведения процесс, в котором учитывается избирание рыбой участков с более высокой плотностью зоопланктона, текущее положение стаи относительно границ области моделирования, направление перемещения на предыдущем временном шаге и время максимального пребывания на заданном участке.
Рыбный косяк рассматривается как некое целостное образование - "super-individual" структура (Scheffer et al., 1995). Он поедает зоопланктон и перемещается по сетке в пределах области интегрирования в соответствии со следующими правилами:
1) рыбная стая может выедать зоопланктон вплоть до установленной минимальной (пороговой) его концентрации, после чего всегда уходит с этого участка;
2) рыбная стая может и должна уйти с участка в том случае, если превышено время максимального пребывания на заданном участке, даже если минимальная (пороговая) концентрация зоопланктона не достигнута;
3) рыбная стая запоминает и предпочитает прежнее направление движения. Новое направление перемещения выбирается случайным образом в пределах заданного "угла видимости" относительно направления на предыдущем шаге по времени.
Показано, что как влияние шума, так и присутствие рыб может индуцировать и ускорять процесс пространственного структурообразования. Таким образом, пятнистость распределения планктона действительно может быть обусловлена как биологическим, так и физическим контролем или же совместным их влиянием. Внесение шума в модель совершенно необходимо для того, чтобы сгенерировать более реалистичную картину пространственного распределения планктона путем сглаживания искусственно возникающих популяционных структур типа спиралей и плоских волн.
Факторы пространственного распределения организмов могут играть важную роль в динамике системы потребитель - ресурс. При одной и той же средней концентрации локальные плотности, как правило, существенно различаются, предопределяя исход биотических взаимоотношений для системы в целом. И, наоборот, при равномерном (то есть без наличия существенных локальных различий) исходном распределении популяций взаимодействия между ними могут приводить к возникновению упорядоченных пространственно-временных структур. В настоящее время при описании структурообразования в биологических системах используется в основном феноменологических подход, опирающийся на обобщенные модели типа адвекция - диффузия. Такой подход можно считать оправданным лишь при анализе макроструктур, охватывающих области распространения отдельных видов или крупных популяций. При рассмотрении же локальных картин пространственного размещения организмов возникает необходимость учета таких факторов структурообразования, как поведенческие реакции отдельных индивидуумов. Эффективным средством изучения в этом случае могут считаться модели иного, механистического типа, например, индивидуально ориентированные.
В работе (Бобырев и др., 2000) рассматривается механизм формирования пространственного размещения в системе потребитель - ресурс, представленной двумя популяциями типа рыба - зоопланктон. Исследование проведено на модели, в явном виде учитывающей характер пространственного распределения организмов. Сценарии модельных экспериментов проигрывались на плоскости размерностью 200?200 ячеек. Внутри популяции хищника индивидуальные различия отсутствуют. В начальный момент времени пространственное распределение отдельных особей хищников задается случайным образом. Мотивацией поведения потребителя выступает требование максимизации скорости потребления корма, что соответствует основным положениям теории оптимального добывания пищи. Потребитель обладает способностью с заданной точностью оценивать плотность кормовых организмов в пределах круговой области. Размеры этой области ограничены дистанцией сенсорного реагирования и представляют собой фиксированный параметр. Перемещения хищника организованы таким образом, что выбор направления движения определяется градиентом плотности корма. Дистанция перемещения потребителя за один временной интервал соответствует расстоянию между смежными ячейками моделируемого пространства. При столкновении потребителя с кормовой частичкой происходит ее поглощение. Считается, что все кормовые организмы характеризуются одинаковыми размерами и уязвимостью для хищника. Смертность корма происходит только за счет выедания. Весовой и линейный рост отсутствует. Начальное распределение корма задается однородным по пространству. На возобновление кормовых организмов наложены пространственные ограничения. На каждом временном шаге фиксированное количество ячеек пространства (их координаты выбираются случайно) приобретают статус "репродуктивных". Прирост происходит только в незанятых ячейках пространства, рядом с которыми находятся "размножающиеся" особи. Таким образом, имеет место анизотропный рост популяции, его темп неявным образом связан с локальной плотностью. Предполагается, что время, затрачиваемое на анализ окружения и поглощение корма, пренебрежимо мало по сравнению со временем, необходимым на перемещение.
В результате моделирования было показано следующее:
1) характерной особенностью поведения модели является формирование устойчивых упорядоченных структур в исходно гомогенном пространственном распределении корма. Конечное распределение корма характеризуется наличием регулярно перемежающихся зон низкой и высокой концентрации кормовых организмов;
2) переходный процесс занимает достаточно длительное время (тысячи модельных временных шагов), в течение которого качественное состояние системы претерпевает ряд последовательных изменений. На начальной стадии имитационного эксперимента траектории движения консументов носят псевдослучайный характер, при отсутствии выраженного приоритетного направления движения. В дальнейшем в результате поглощения консументами и возобновления пищевых частиц в распределении корма образуются области (узлы) пониженной концентрации корма (выражена сетчатая пространственная структура). С течением времени узлы соединяются между собой, формируя четкую структуру, дальнейшая эволюция которой приводит к образованию стабильного пространственного паттерна, представленного чередующимися полосами. Стабилизация пространственной структуры сопровождается стабилизацией плотности кормовых организмов и среднего рациона потребителей;
3) из общих соображений можно заключить, что процесс возникновения структуры будет определяться соотношением количества хищников (их суммарным пищевым рационом) и скоростью воспроизводства популяции кормовых организмов. При малом количестве хищников и быстром заполнении свободного пространства кормовыми частичками структура не возникает. При фиксированных параметрах воспроизводства корма увеличение числа потребителей имеет следствием образование структуры, причем скорость ее образования тем выше, чем больше число потребителей;
4) устойчивость формирующейся зональности в размещении кормовых организмов обеспечивается принятой в модели системой ориентации консументов. Ее следствием является стремление потребителя избегать участков пространства, содержащих следы предшествующей деятельности, как собственной, так и других потребителей (в пределах области реагирования). Это приводит к тому, что в распределении ресурса образуются области "разрежения" (узлы), отделенные друг от друга расстоянием, приблизительно равным дистанции сенсорного реагирования;
5) стохастические компоненты поведения (ошибочное определение плотности кормовых частиц, случайные изменения направления движений и т. п.), в целом замедляя процессы формирования структур, качественно не меняют поведения модели;
6) в модели не задана в явном виде какая-либо конкуренция, но сама система ориентации порождает разделение пространства консументами и определенную регулярность их положения.
Многие традиционные исследования пространственной неоднородности рассматривают динамику отдельно взятой популяции, распределенной по участкам в окружающем их пространстве. В таких моделях скорости распространения достаточно высоки, а перемещение может зависеть от плотности. Более современные подходы придают особое значение сохранению ансамбля, состоящего из многих локальных популяций, то есть сохранению метапопуляции. В метапопуляционных моделях ее распространение, как правило, не зависит от плотности, а скорости перемещения между локальными популяциями низки. Используя эти положения, Тейлор (Taylor, 1991) рассмотрел вопрос о том, действительно ли многие природные взаимодействия по типу "хищник-жертва" могут быть описаны как истинные метапопуляции, где множество локально неустойчивых популяций сохраняются в результате распространения. Он пришел к выводу, что хотя по существу найдется очень немного примеров метапопуляций в нашем понимании этого термина, тем не менее многие системы на самом деле могут объединять в себе характерные особенности как отдельных популяций, так и метапопуляций. Поэтому трудно провести четкое различие между этими двумя пространственными масштабами и соответствующими им уровнями описания. В работе (Rohani, Miramontes, 1995) рассматривается один из примеров таких смешанных структур, где отдельно взятая популяция паразитов захватывает популяцию хозяина. Мы рассматриваем эту модель специально для того, чтобы показать, как чисто биологические эффекты, без учета адвекции и диффузии, могут приводить к появлению пространственных структур.
Используется разработанная ранее модель "паразит-хозяин" (Hassell et al., 1991; Comins et al., 1992). С помощью этой модели показано, что (как и предсказывает теория) объединение некоторого количества неустойчивых популяций посредством их локального перемещения действительно способствует сохранению метапопуляции. Более того, было показано, что в больших областях параметрического пространства взаимодействия по типу "паразит - хозяин" способны порождать упорядоченные пространственные структуры, например такие, как спирали Обратим внимание на то, что здесь нет прямого воздействия адвекции и диффузии, как в океанах или морях. Это является лишним подтверждением важности именно трофических (или каких-либо еще) нелинейных взаимодействий в самой биологической системе при образовании пространственных неоднородностей. В исходной модели предполагается, что между двумя последовательными генерациями, постоянная доля паразитов и их хозяев мигрирует и колонизирует в равных пропорциях ближайшие соседние восемь участков. Этот процесс распространения может зависеть от плотности так, что большее количество хозяев (и меньшее количество паразитов) мигрирует из участков с большей их плотностью, однако вероятность того, что они окажутся на одном из соседних участков одинакова. В данной работе рассматривается еще один возможный модельный сценарий: распространение хозяев единообразно, в то время как паразиты предпочитают перемещаться в те области, где вероятность встречи с еще незараженными особями более высока. В рассматриваемой модели генерации паразитов и хозяев дискретны и описываются разностными уравнениями. Распространение происходит локально, в пределах пространственно структурированной окружающей среды.
Модель по своей структуре очень проста. Рассматривается среда, которая разделена на отдельные дискретные двумерные участки. На этих участках распределены паразиты и хозяева. Каждый шаг модели (каждая генерация) состоит их двух различных фаз:
1) фазы распространения популяций;
2) фазы популяционных отношений и межвидовой динамики (репродукция и паразитизм).
В течение первой фазы некоторая доля хозяев ![]() покидает свой текущий участок, в то время как оставшиеся особи размножаются на этом же участке. Распространяющиеся особи занимают в равных пропорциях ближайшие восемь участков в соответствии со следующим уравнением:
покидает свой текущий участок, в то время как оставшиеся особи размножаются на этом же участке. Распространяющиеся особи занимают в равных пропорциях ближайшие восемь участков в соответствии со следующим уравнением:
![]() , (47)
, (47)
где ![]() и
и ![]() представляют собой плотности хозяев перед и после распространения на каком-либо из участков для генерации t.
представляют собой плотности хозяев перед и после распространения на каком-либо из участков для генерации t. ![]() - средняя плотность популяции, полученная осреднением по ближайшим восьми участкам, непосредственно примыкающих к данному. В отличие от прежних моделей (Hassell et al., 1991; Solй et al., 1992) правило, согласно которому происходит распространение популяции паразитов, более сложное: вместо равновероятного перемещения к любому из ближайших соседних участков в данной модели задается определенное предпочтение и паразиты с большей вероятностью перемещаются на те участки, где имеется более высокая плотность хозяев. Для того, чтобы задать это предпочтение, используется индекс агрегированности
- средняя плотность популяции, полученная осреднением по ближайшим восьми участкам, непосредственно примыкающих к данному. В отличие от прежних моделей (Hassell et al., 1991; Solй et al., 1992) правило, согласно которому происходит распространение популяции паразитов, более сложное: вместо равновероятного перемещения к любому из ближайших соседних участков в данной модели задается определенное предпочтение и паразиты с большей вероятностью перемещаются на те участки, где имеется более высокая плотность хозяев. Для того, чтобы задать это предпочтение, используется индекс агрегированности ![]() (Hassell, May, 1973), а доля
(Hassell, May, 1973), а доля ![]() распространяющихся паразитов, попадающих на новый участок, определяется следующим соотношением:
распространяющихся паразитов, попадающих на новый участок, определяется следующим соотношением: ![]() .
. ![]() - общее количество (общая плотность) хозяев на новом участке и ближайших к нему восьми участках, k - нормировочный множитель, который используется для того, чтобы выразить
- общее количество (общая плотность) хозяев на новом участке и ближайших к нему восьми участках, k - нормировочный множитель, который используется для того, чтобы выразить ![]() в долях единицы. Когда индекс агрегированности
в долях единицы. Когда индекс агрегированности ![]() , распространение паразитов происходит в любом направлении одинаково и не зависит от плотности. Если
, распространение паразитов происходит в любом направлении одинаково и не зависит от плотности. Если ![]() , то паразиты заселяют участки в одинаковой пропорции по отношению к доле хозяев. Если же
, то паразиты заселяют участки в одинаковой пропорции по отношению к доле хозяев. Если же ![]() , то паразиты демонстрируют различное предпочтение участков, содержащих наибольшие плотности хозяев. Таким образом, уравнение, описывающее распространение паразитов, имеет следующий вид:
, то паразиты демонстрируют различное предпочтение участков, содержащих наибольшие плотности хозяев. Таким образом, уравнение, описывающее распространение паразитов, имеет следующий вид:
![]() , (48)
, (48)
где ![]() и
и ![]() - плотности паразитов для генерации t на каком-либо участке до и после распространения,
- плотности паразитов для генерации t на каком-либо участке до и после распространения, ![]() - доля взрослых самок паразитов. В модели использовался вариант с поглощающими граничными условиями, хотя, как показали результаты моделирования, и при других вариантах граничных условий результаты, по крайней мере качественно, совпадают. Фаза репродукции и паразитизма описывается в модели в соответствии с параметризацией этих процессов, предложенных Николсоном и Бейли (Nicholson, Bailey, 1935). В соответствии с этим предполагается, что все участки качественно одинаковы, а паразитизм происходит случайным образом. В таком случае:
- доля взрослых самок паразитов. В модели использовался вариант с поглощающими граничными условиями, хотя, как показали результаты моделирования, и при других вариантах граничных условий результаты, по крайней мере качественно, совпадают. Фаза репродукции и паразитизма описывается в модели в соответствии с параметризацией этих процессов, предложенных Николсоном и Бейли (Nicholson, Bailey, 1935). В соответствии с этим предполагается, что все участки качественно одинаковы, а паразитизм происходит случайным образом. В таком случае:
![]() . (49)
. (49)
Здесь ![]() - скорость роста популяции хозяев, а - удельная скорость атак и b - коэффициент эффективности превращения зараженных хозяев во взрослых самок паразитов на следующей генерации.
- скорость роста популяции хозяев, а - удельная скорость атак и b - коэффициент эффективности превращения зараженных хозяев во взрослых самок паразитов на следующей генерации.
Путем численных экспериментов с данной моделью показано, что агрегирование паразитов на участках с высокими плотностями хозяев способствует самоорганизации и образованию спиральных волн. Тем не менее, при высоких скоростях распространения паразитов значительные уровни агрегирования препятствуют образованию спиралевидных структур. Это объясняется тем, что при ![]() большинство распространяющихся паразитов переходит на участки, содержащие поблизости наибольшие плотности хозяев. А поскольку участки высокой плотности формируют основную часть спиралей, повышенное агрегирование подавляет образование спиральных структур. Таким образом, на пространственную динамику большое внимание оказывает значение коэффициента агрегированности. Было установлено, что при
большинство распространяющихся паразитов переходит на участки, содержащие поблизости наибольшие плотности хозяев. А поскольку участки высокой плотности формируют основную часть спиралей, повышенное агрегирование подавляет образование спиральных структур. Таким образом, на пространственную динамику большое внимание оказывает значение коэффициента агрегированности. Было установлено, что при ![]() никакой упорядоченности не наблюдается. Затем, по мере роста
никакой упорядоченности не наблюдается. Затем, по мере роста ![]() , появляются спирали сначала небольшого размера, потом они становятся все более отчетливо выраженными. Высокие уровни агрегированности вновь постепенно подавляют процесс образования спиралей, в результате чего пространственная динамика становится слабо упорядоченной. Было также показано, что изменчивость плотностей популяций тесно коррелирует со степенью пространственной упорядоченности в системе: области с некоторым количеством больших, хорошо выраженных спиралей, обладали наименьшей изменчивостью популяционной плотности. При рассмотрении средних плотностей популяции хозяев выяснились интересные особенности: эти плотности были наименьшими в том случае, когда пространственная динамика состояла их большого количества мелких спиралей на фоне временно неупорядоченных структур. Таким образом, эффективность поиска у паразитов невелика в тех случаях, когда пространство характеризуется или наличием крупных спиралей или отсутствием упорядоченности. Наоборот, эффективность поиска наивысшая тогда, когда хозяева формируют мелкие спиральные структуры, то есть паразиты используют среду наилучшим для себя образом в том случае, если имеется некоторое смешение пространственной упорядоченности и беспорядка.
, появляются спирали сначала небольшого размера, потом они становятся все более отчетливо выраженными. Высокие уровни агрегированности вновь постепенно подавляют процесс образования спиралей, в результате чего пространственная динамика становится слабо упорядоченной. Было также показано, что изменчивость плотностей популяций тесно коррелирует со степенью пространственной упорядоченности в системе: области с некоторым количеством больших, хорошо выраженных спиралей, обладали наименьшей изменчивостью популяционной плотности. При рассмотрении средних плотностей популяции хозяев выяснились интересные особенности: эти плотности были наименьшими в том случае, когда пространственная динамика состояла их большого количества мелких спиралей на фоне временно неупорядоченных структур. Таким образом, эффективность поиска у паразитов невелика в тех случаях, когда пространство характеризуется или наличием крупных спиралей или отсутствием упорядоченности. Наоборот, эффективность поиска наивысшая тогда, когда хозяева формируют мелкие спиральные структуры, то есть паразиты используют среду наилучшим для себя образом в том случае, если имеется некоторое смешение пространственной упорядоченности и беспорядка.
В работе (Solй et al., 1992) рассматривается двумерная CML-модель (Coupled Map Lattice), представляющая собой систему нелинейных разностных уравнений:
![]() . (50)
. (50)
Здесь ![]() - популяции жертвы и хищника соответственно. Предполагается, что
- популяции жертвы и хищника соответственно. Предполагается, что ![]() и U является открытым множеством. Кроме того, для функции
и U является открытым множеством. Кроме того, для функции ![]() вводятся еще два ограничения:
вводятся еще два ограничения: ![]() . В данной статье
. В данной статье ![]() рассматривается в виде
рассматривается в виде ![]() . При исследовании системы (50) использовали постоянное значение параметра роста
. При исследовании системы (50) использовали постоянное значение параметра роста ![]() и несколько комбинаций параметра
и несколько комбинаций параметра ![]() . Показано, что для
. Показано, что для ![]() в системе присутствует хаотическая динамика как следствие доминирующей роли логистической части, то есть множителя
в системе присутствует хаотическая динамика как следствие доминирующей роли логистической части, то есть множителя ![]() . Для
. Для ![]() хаотический аттрактор сначала разрушается и в системе устанавливается устойчивое состояние, которое затем последовательно меняется на квазипериодический аттрактор и постепенно, с ростом параметра
хаотический аттрактор сначала разрушается и в системе устанавливается устойчивое состояние, которое затем последовательно меняется на квазипериодический аттрактор и постепенно, с ростом параметра ![]() , в системе снова устанавливается хаотический режим.
, в системе снова устанавливается хаотический режим.
Модель можно расширить и учесть влияние диффузии:
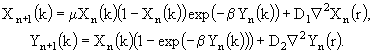 (51)
(51)
В (51) дискретный оператор диффузии задается следующими соотношениями: 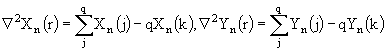 . При расчетах принято значение q, равное 4. Кроме того, для того чтобы избежать появления отрицательных значений плотности популяции, задавалось условие: если
. При расчетах принято значение q, равное 4. Кроме того, для того чтобы избежать появления отрицательных значений плотности популяции, задавалось условие: если ![]() , то
, то ![]() . Граничные условия: либо периодические, либо задавалось условие отсутствия потоков через границу. Размер расчетной области: 512?512 ячеек. Устойчивые спиральные волны наблюдались вплоть до
. Граничные условия: либо периодические, либо задавалось условие отсутствия потоков через границу. Размер расчетной области: 512?512 ячеек. Устойчивые спиральные волны наблюдались вплоть до ![]() , при еще больших значениях этого параметра образование структур не наблюдалось. Хаотические спирали при больших
, при еще больших значениях этого параметра образование структур не наблюдалось. Хаотические спирали при больших ![]() локально неустойчивы. Они перемещаются по сетке, создаются и разрушаются. Текущая позиция данной спирали изменяется со временем, по- видимому, случайным образом.
локально неустойчивы. Они перемещаются по сетке, создаются и разрушаются. Текущая позиция данной спирали изменяется со временем, по- видимому, случайным образом.
В работе (Петровский и др., 1998) исследуется еще один важный сценарий формирования неоднородностей в планктонном сообществе: образование пространственных структур в нелинейных диссипативных системах (в частности, типа хищник - жертва) при инвазии популяций и одинаковых коэффициентах диффузии хищника и жертвы. Рассмотрим динамику двухкомпонентной активной среды, описываемой системой уравнений с нелинейностью типа "хищник - жертва":
![]() , (52)
, (52)
![]() . (53)
. (53)
Функция ![]() обладает следующими свойствами:
обладает следующими свойствами: ![]() ,
, ![]() ;
; ![]() ;
; ![]() ,
, ![]() . Выберем функцию
. Выберем функцию ![]() следующего вида (для модели с небольшим числом переменных поиск точного вида зависимости не имеет смысла, достаточно потребовать, чтобы она описывала принципиальные особенности локальной динамики):
следующего вида (для модели с небольшим числом переменных поиск точного вида зависимости не имеет смысла, достаточно потребовать, чтобы она описывала принципиальные особенности локальной динамики):
![]() , (54)
, (54)
где a и b - некоторые постоянные, зависящие от характеристик окружающей среды. Параметр b имеет смысл "емкости" среды для данной популяции. Процесс выедания ![]() описывается модифицированной функцией Ивлева:
описывается модифицированной функцией Ивлева:
![]() , (55)
, (55)
где I, ![]() ,
, ![]() - постоянные. При концентрации жертвы меньше порогового значения
- постоянные. При концентрации жертвы меньше порогового значения ![]() питание хищника прекращается. Величина параметра
питание хищника прекращается. Величина параметра ![]() может быть различной для разных видов. Учитывая, что, как правило,
может быть различной для разных видов. Учитывая, что, как правило, ![]() , в данной работе принято, что
, в данной работе принято, что ![]() . Для дальнейшего исследования системы уравнений (52) - (55) введем безразмерные переменные:
. Для дальнейшего исследования системы уравнений (52) - (55) введем безразмерные переменные: ![]() . В новых переменных система принимает вид (штрихи далее опущены):
. В новых переменных система принимает вид (штрихи далее опущены):
![]() , (56)
, (56)
![]() , (57)
, (57)
где ![]() - безразмерные параметры. Начальные условия для уравнений (56), (57) используем в следующем виде:
- безразмерные параметры. Начальные условия для уравнений (56), (57) используем в следующем виде:
![]() , (58)
, (58)
![]() . (59)
. (59)
Здесь безразмерные параметры ![]() имеют смысл, соответственно, ширины и амплитуды начальных популяционных вспышек для хищника и жертвы.
имеют смысл, соответственно, ширины и амплитуды начальных популяционных вспышек для хищника и жертвы.
Численное исследование данной системы уравнений показало, что инвазия популяций от места их первоначального вселения происходит в виде движения фронтов - "популяционных волн". Эти фронты, в зависимости от параметров сообщества, распространяются либо вместе, либо поочередно, и могут иметь достаточно сложную осциллирующую структуру. При этом осцилляции фронта могут быть столь существенны, что распространение популяций фактически происходит в виде движения отдельных пятен. Также показано, что при некоторых значениях параметров после прохождения популяционных волн в системе устанавливаются нерегулярные пространственно-временные колебания, имеющие вид эволюционирующей "пятнистой" структуры. Образование пространственных структур в нелинейных диссипативных системах (в частности, типа хищник - жертва) при различающихся коэффициентах диффузии ![]() - хорошо известный факт (Свирежев, 1987). Возникновение же структур в системе такого типа при
- хорошо известный факт (Свирежев, 1987). Возникновение же структур в системе такого типа при ![]() - новый результат, полученный впервые в данной работе.
- новый результат, полученный впервые в данной работе.
Дополнительная информация:
© 2001-2026 Кафедра биофизики МГУ