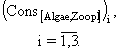Рассмотрим основные принципы, на основании которых были построены несколько имитационных моделей водных экосистем, рассматривающих процессы трансформации соединений азота или фосфора.
При создании математической модели круговорота азота в пресноводных экосистемах в работе (Добрынский, Рогаль, 1997) выделяли следующие укрупненные компоненты экосистемы: 1) водные растворы неорганических соединений азота (аммонийный, нитратный и нитритный); 2) водорастворимые органические формы азота; 3) взвеси водонерастворимых веществ типа детрита; 4) фитопланктон; 5) зоопланктон; 6) бактериопланктон; 7) бентос; 8) высшая водная растительность; 9) ихтиофауна. Если исключить процессы пополнения и убыли азота в водоемах за счет притока и стока поверхностных вод, то остальные превращения азота практически полностью определяются деятельностью микроорганизмов. При этом все процессы, связанные с круговоротом азота, можно разбить на три группы: 1) процессы фиксации свободного азота; 2) процессы превращения одних форм азота в другие; 3) процессы, ведущие к обеднению водоема связанным азотом (Горленко и др., 1977). Фиксацию атмосферного азота осуществляет широкий спектр микроорганизмов, в частности водородокисляющие бактерии м синезеленые водоросли. Наблюдается прямая корреляция между количеством фиксированного азота и суммарным развитием (биомассой) синезеленых водорослей. При этом скорость фиксации азота на свету примерно в 10 раз больше, чем в темноте и совпадает с интенсивностью фотосинтеза. Эффективность бактериальной фиксации азота заметно возрастает при низкой концентрации его органических соединений в водоеме и при пониженном парциальном давлении кислорода. При минерализации органический азот превращается в аммиак. Процесс минерализации отмершего планктона начинается в водной массе и продолжается в поверхностном слое ила. Процессы, противоположные азотфиксации, могут протекать в водоемах по-разному. При прямой денитрификации восстановление нитратов идет до свободного азота. Однако лишь немногие виды являются истинными денитрификаторами. Большинство же процессов денитрификации протекает в несколько стадий. В результате нитраты вначале восстанавливаются до аммиака или закиси азота, а затем до свободного азота (Горленко и др., 1977) При построении математической модели предполагается, что влиянием зоопланктона, зообентоса и ихтиофауны на внутриводоемные процессы трансформации вещества можно пренебречь. В данной работе разные неорганические соединения азота не разделяются на отдельные компоненты. Участие высшей водной растительности во внутриводоемных процессах представлено в модели как действие некоторого вполне определенного по величине внешнего фактора. Каждая компонента азотной системы характеризуется свой концентрацией. Ее величина для таких компонент, как азот водорослей, бактериальный, детрита и макрофитов измеряется отношением общего количества азота в сырой массе вещества каждой из указанных компонент к тому объему воды, в котором они рассматриваются. Основные гипотезы (предположения), на основании которых производится вывод уравнений, описывающих круговорот азота в пресноводных экосистемах, состоят в следующем:
1. Компоненты азотной системы равномерно распределены по объему водоема, то есть их концентрация не зависит от пространственных координат.
2. Концентрация свободного азота ![]() в водоеме пропорциональна парциальному атмосферному давлению воздуха, то есть не зависит от протекающих в водоеме биохимических процессов трансформации соединений азота.
в водоеме пропорциональна парциальному атмосферному давлению воздуха, то есть не зависит от протекающих в водоеме биохимических процессов трансформации соединений азота.
3. Бактерии-азотфиксаторы ![]() поглощают свободный (атмосферный) азот
поглощают свободный (атмосферный) азот ![]() в количестве, пропорциональном их биомассе, то есть происходит трансформация свободного азота в азот биомассы бактерий-азотфиксаторов
в количестве, пропорциональном их биомассе, то есть происходит трансформация свободного азота в азот биомассы бактерий-азотфиксаторов ![]() . Коэффициент пропорциональности (поглощения) зависит от концентрации органического азота, легкоусвояемого органического вещества и температуры воды.
. Коэффициент пропорциональности (поглощения) зависит от концентрации органического азота, легкоусвояемого органического вещества и температуры воды.
4. Водоросли ![]() , поглощая свободный азот
, поглощая свободный азот ![]() , превращают его в свою биомассу
, превращают его в свою биомассу ![]() . Коэффициент пропорциональности зависит от концентрации неорганического азота, интенсивности солнечной радиации, температуры воды, освещенности и других факторов.
. Коэффициент пропорциональности зависит от концентрации неорганического азота, интенсивности солнечной радиации, температуры воды, освещенности и других факторов.
5. Бактерии-азотфиксаторы ![]() вместе с продуктами метаболизма выделяют водорастворимые органические соединения азота
вместе с продуктами метаболизма выделяют водорастворимые органические соединения азота ![]() , увеличивая при этом общую массу органического азота в количестве, пропорциональном биомассе бактерий, то есть происходит трансформация азота бактерий-азотфиксаторов в органический азот
, увеличивая при этом общую массу органического азота в количестве, пропорциональном биомассе бактерий, то есть происходит трансформация азота бактерий-азотфиксаторов в органический азот ![]() . Коэффициент пропорциональности зависит от температуры воды и концентрации легкоусвояемого органического вещества.
. Коэффициент пропорциональности зависит от температуры воды и концентрации легкоусвояемого органического вещества.
6. Бактерии-азотфиксаторы ![]() , отмирая, пополняют (увеличивают) массу азота детрита
, отмирая, пополняют (увеличивают) массу азота детрита ![]() в количестве, пропорциональном биомассе бактерий, то есть происходит трансформация азота бактерий-азотфиксаторов в азот детрита
в количестве, пропорциональном биомассе бактерий, то есть происходит трансформация азота бактерий-азотфиксаторов в азот детрита ![]() . Коэффициент пропорциональности зависит от температуры воды, кислорода и других факторов.
. Коэффициент пропорциональности зависит от температуры воды, кислорода и других факторов.
7. Бактерии-деазотфиксаторы ![]() выделяют свободный азот
выделяют свободный азот ![]() в количестве, пропорциональном их биомассе, то есть происходит трансформация азота бактерий-деазотфиксаторов в свободный азот
в количестве, пропорциональном их биомассе, то есть происходит трансформация азота бактерий-деазотфиксаторов в свободный азот ![]() . Коэффициент пропорциональности зависит от легкоусвояемого органического вещества, концентрации неорганического азота, температуры воды и наличия кислорода (этот процесс происходит более активно при низкой концентрации кислорода).
. Коэффициент пропорциональности зависит от легкоусвояемого органического вещества, концентрации неорганического азота, температуры воды и наличия кислорода (этот процесс происходит более активно при низкой концентрации кислорода).
8. Бактерии-деазотфиксаторы ![]() , отмирая, пополняют (увеличивают) массу азота детрита
, отмирая, пополняют (увеличивают) массу азота детрита ![]() в количестве, пропорциональном биомассе бактерий, то есть происходит трансформация азота бактерий-деазотфиксаторов в азот детрита
в количестве, пропорциональном биомассе бактерий, то есть происходит трансформация азота бактерий-деазотфиксаторов в азот детрита ![]() . Коэффициент пропорциональности зависит от концентрации неорганического азота, температуры воды и кислорода.
. Коэффициент пропорциональности зависит от концентрации неорганического азота, температуры воды и кислорода.
9. Бактерии-минерализаторы ![]() с продуктами метаболизма выделяют неорганический азот
с продуктами метаболизма выделяют неорганический азот ![]() в количестве, пропорциональном их биомассе, то есть происходит трансформация азота бактерий-минерализаторов в неорганический азот
в количестве, пропорциональном их биомассе, то есть происходит трансформация азота бактерий-минерализаторов в неорганический азот ![]() . Коэффициент пропорциональности зависит от температуры воды, наличия кислорода, легкоусвояемого органического вещества и концентрации органического азота.
. Коэффициент пропорциональности зависит от температуры воды, наличия кислорода, легкоусвояемого органического вещества и концентрации органического азота.
10.Бактерии-минерализаторы ![]() , отмирая, пополняют (увеличивают) массу азота детрита
, отмирая, пополняют (увеличивают) массу азота детрита ![]() в количестве, пропорциональном биомассе бактерий, то есть происходит трансформация азота бактерий-деазотфиксаторов в азот детрита
в количестве, пропорциональном биомассе бактерий, то есть происходит трансформация азота бактерий-деазотфиксаторов в азот детрита ![]() . Коэффициент пропорциональности зависит от концентрации кислорода и температуры воды.
. Коэффициент пропорциональности зависит от концентрации кислорода и температуры воды.
11.Бактерии-деазотфиксаторы ![]() , поглощая неорганический азот
, поглощая неорганический азот ![]() , увеличивают свою биомассу в количестве, пропорциональном их биомассе и концентрации неорганического азота, то есть происходит трансформация неорганического азота в азот бактерий-деазотфиксаторов
, увеличивают свою биомассу в количестве, пропорциональном их биомассе и концентрации неорганического азота, то есть происходит трансформация неорганического азота в азот бактерий-деазотфиксаторов ![]() . Коэффициент пропорциональности зависит от концентрации неорганического азота, наличия легкоусвояемых органических веществ, концентрации кислорода и температуры воды.
. Коэффициент пропорциональности зависит от концентрации неорганического азота, наличия легкоусвояемых органических веществ, концентрации кислорода и температуры воды.
12.Водоросли ![]() , поглощая неорганический азот
, поглощая неорганический азот ![]() , превращают его в свою биомассу в количестве, пропорциональном его биомассе и концентрации неорганического азота, то есть происходит трансформация неорганического азота в азот фитопланктона
, превращают его в свою биомассу в количестве, пропорциональном его биомассе и концентрации неорганического азота, то есть происходит трансформация неорганического азота в азот фитопланктона ![]() . Коэффициент пропорциональности зависит от концентрации неорганического азота, интенсивности солнечной радиации, температуры воды, освещенности, наличия легкоусвояемого органического вещества и других факторов.
. Коэффициент пропорциональности зависит от концентрации неорганического азота, интенсивности солнечной радиации, температуры воды, освещенности, наличия легкоусвояемого органического вещества и других факторов.
13.Высшая водная растительность ![]() , потребляя неорганический азот
, потребляя неорганический азот ![]() , увеличивает свою биомассу, то есть происходит трансформация неорганического азота в азот высшей водной растительности в количестве, пропорциональном биомассе высшей водной растительности и концентрации неорганического азота
, увеличивает свою биомассу, то есть происходит трансформация неорганического азота в азот высшей водной растительности в количестве, пропорциональном биомассе высшей водной растительности и концентрации неорганического азота ![]() . Коэффициент пропорциональности зависит от концентрации неорганического азота в донных отложениях, температуры воды, интенсивности солнечной радиации, условий освещенности и других факторов.
. Коэффициент пропорциональности зависит от концентрации неорганического азота в донных отложениях, температуры воды, интенсивности солнечной радиации, условий освещенности и других факторов.
14.Бактерии-минерализаторы ![]() , потребляя органический азот
, потребляя органический азот ![]() , увеличивают свою биомассу в количестве, пропорциональном их биомассе и концентрации органического азота, то есть происходит трансформация органического азота в азот бактерий-минерализаторов
, увеличивают свою биомассу в количестве, пропорциональном их биомассе и концентрации органического азота, то есть происходит трансформация органического азота в азот бактерий-минерализаторов ![]() . Коэффициент пропорциональности зависит от концентрации органического азота, наличия легкоусвояемого органического вещества, температуры воды и других факторов.
. Коэффициент пропорциональности зависит от концентрации органического азота, наличия легкоусвояемого органического вещества, температуры воды и других факторов.
15.Водоросли ![]() в процессе жизнедеятельности с продуктами метаболизма выделяют соединения органического азота
в процессе жизнедеятельности с продуктами метаболизма выделяют соединения органического азота ![]() в количестве, пропорциональном их биомассе и концентрации неорганического азота, то есть происходит трансформация азота водорослей в органический азот
в количестве, пропорциональном их биомассе и концентрации неорганического азота, то есть происходит трансформация азота водорослей в органический азот ![]() . Коэффициент пропорциональности зависит от концентрации неорганического азота, интенсивности солнечной радиации, условий освещенности и температуры воды.
. Коэффициент пропорциональности зависит от концентрации неорганического азота, интенсивности солнечной радиации, условий освещенности и температуры воды.
16.Водоросли ![]() , отмирая, увеличивают массу азота детрита
, отмирая, увеличивают массу азота детрита ![]() в количестве, пропорциональном их биомассе, то есть происходит трансформация азота водорослей в азот детрита
в количестве, пропорциональном их биомассе, то есть происходит трансформация азота водорослей в азот детрита ![]() . Коэффициент пропорциональности зависит от концентрации неорганического азота, интенсивности солнечной радиации, условий освещенности и температуры воды.
. Коэффициент пропорциональности зависит от концентрации неорганического азота, интенсивности солнечной радиации, условий освещенности и температуры воды.
17.Бактерии-минерализаторы ![]() , потребляя детрит
, потребляя детрит ![]() , увеличивают свою биомассу в количестве, пропорциональном их биомассе и концентрации азота детрита, то есть происходит трансформация азота детрита в азот бактерий-минерализаторов
, увеличивают свою биомассу в количестве, пропорциональном их биомассе и концентрации азота детрита, то есть происходит трансформация азота детрита в азот бактерий-минерализаторов ![]() . Коэффициент пропорциональности зависит от концентрации органического азота, наличия легкоусвояемых органических веществ, температуры воды и концентрации кислорода в воде.
. Коэффициент пропорциональности зависит от концентрации органического азота, наличия легкоусвояемых органических веществ, температуры воды и концентрации кислорода в воде.
18.Детрит ![]() седиментирует в донные отложения в количестве, пропорциональном его массе, то есть происходит трансформация азота детрита в азот донных отложений.
седиментирует в донные отложения в количестве, пропорциональном его массе, то есть происходит трансформация азота детрита в азот донных отложений.
19.Высшая водная растительность ![]() в процессе жизнедеятельности с продуктами метаболизма выделяет соединения органического азота
в процессе жизнедеятельности с продуктами метаболизма выделяет соединения органического азота ![]() в количестве, пропорциональном ее биомассе, то есть происходит трансформация азота высшей водной растительности в органический азот
в количестве, пропорциональном ее биомассе, то есть происходит трансформация азота высшей водной растительности в органический азот ![]() . Коэффициент пропорциональности зависит от температуры воды, интенсивности солнечной радиации, условий освещенности и других факторов.
. Коэффициент пропорциональности зависит от температуры воды, интенсивности солнечной радиации, условий освещенности и других факторов.
20.Отмирая, высшая водная растительность ![]() пополняет массу азота детрита
пополняет массу азота детрита ![]() в количестве, пропорциональном ее биомассе, то есть происходит трансформация азота макрофитов в азот детрита
в количестве, пропорциональном ее биомассе, то есть происходит трансформация азота макрофитов в азот детрита ![]() . Коэффициент пропорциональности зависит от температуры воды, интенсивности солнечной радиации и условий освещенности.
. Коэффициент пропорциональности зависит от температуры воды, интенсивности солнечной радиации и условий освещенности.
В данной модели учтено влияние кислородного режима на соответствующие коэффициенты и процессы трансформации веществ.
Таким образом, предполагается, что основными переменными модели круговорота азота в водных экосистемах являются следующие величины:
![]()
![]() - концентрация (масса) свободного азота;
- концентрация (масса) свободного азота;
![]()
![]() - концентрация (масса) азота в бактериях-азотфиксаторах (мг/л);
- концентрация (масса) азота в бактериях-азотфиксаторах (мг/л);
![]()
![]() - концентрация (масса) азота в бактериях-денитрификаторах (мг/л);
- концентрация (масса) азота в бактериях-денитрификаторах (мг/л);
![]()
![]() - концентрация (масса) азота в бактериях-минерализаторах (мг/л);
- концентрация (масса) азота в бактериях-минерализаторах (мг/л);
![]()
![]() - концентрация (масса) неорганического азота в растворе (мг/л);
- концентрация (масса) неорганического азота в растворе (мг/л);
![]()
![]() - концентрация (масса) органического азота в растворе (мг/л);
- концентрация (масса) органического азота в растворе (мг/л);
![]()
![]() - концентрация (масса) азота в водорослях (мг/л);
- концентрация (масса) азота в водорослях (мг/л);
![]()
![]() - концентрация (масса) детритного азота (мг/л);
- концентрация (масса) детритного азота (мг/л);
![]()
![]() - концентрация (масса) азота высшей водной растительности (мг/л);
- концентрация (масса) азота высшей водной растительности (мг/л);
W - текущий объем водоема.
Перейдем теперь к выводу уравнений модели. Согласно приведенным выше гипотезам, имеем:
уравнение изменения массы азота бактерий-азотфиксаторов -
![]() (1)
(1)
уравнение изменения массы азота бактерий-денитрификаторов -
![]() (2)
(2)
уравнение изменения массы азота бактерий-минерализаторов -
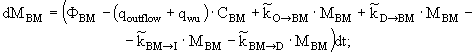 (3)
(3)
уравнение изменения массы неорганических форм азота -
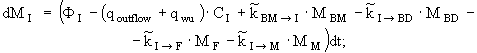 (4)
(4)
уравнение изменения массы водорастворимых органических форм азота -
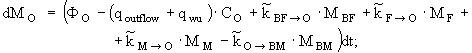 (5)
(5)
уравнение изменения массы азота водорослей -
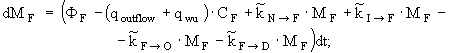 (6)
(6)
уравнение изменения массы детритного азота -
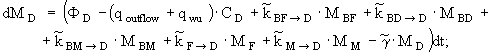 (7)
(7)
уравнение изменения массы азота высшей водной растительности -
![]() . (8)
. (8)
Здесь ![]() , где i = BF, BD, BM, I, O, F, D, M - полное приращение i-ой формы азота за интервал времени dt;
, где i = BF, BD, BM, I, O, F, D, M - полное приращение i-ой формы азота за интервал времени dt; ![]() - скорость поступления i-ой формы азота в экосистему из внешних источников; выражение вида
- скорость поступления i-ой формы азота в экосистему из внешних источников; выражение вида ![]() учитывает изменение i-ой компоненты азотной системы вследствие стока воды из водоема и водопотребления;
учитывает изменение i-ой компоненты азотной системы вследствие стока воды из водоема и водопотребления; ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
,![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() - эмпирические коэффициенты, которые сложным образом зависят от факторов внешней среды и учитывают внутренние механизмы трансформации соединений азота водорослями, бактериями и высшей водной растительностью;
- эмпирические коэффициенты, которые сложным образом зависят от факторов внешней среды и учитывают внутренние механизмы трансформации соединений азота водорослями, бактериями и высшей водной растительностью; ![]() - коэффициент седиментации. При недостаточно полной информации о биохимических процессах, протекающих в пресноводных экосистемах, используются простейшие зависимости коэффициентов от концентраций веществ.
- коэффициент седиментации. При недостаточно полной информации о биохимических процессах, протекающих в пресноводных экосистемах, используются простейшие зависимости коэффициентов от концентраций веществ.
Возвращаясь к уравнениям модели, заметим, что согласно гипотезе об изотропности распределения каждой из компонент азотной системы по водоему имеет место равенство
![]() , i = BF, BD, BM, I, O, F, D, M, (9)
, i = BF, BD, BM, I, O, F, D, M, (9)
где ![]() - текущий объем воды в озере, который определяется из уравнения водного баланса. Учитывая, что
- текущий объем воды в озере, который определяется из уравнения водного баланса. Учитывая, что ![]() , и подставляя вышеуказанные выражения в уравнения (1) - (8) и деля на dt, окончательно получаем следующую систему уравнений для неизвестных концентраций
, и подставляя вышеуказанные выражения в уравнения (1) - (8) и деля на dt, окончательно получаем следующую систему уравнений для неизвестных концентраций ![]() :
:
![]() (10)
(10)
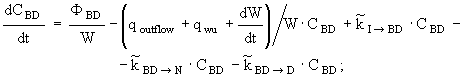 (11)
(11)
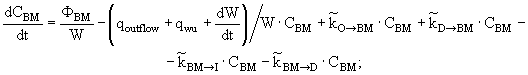 (12)
(12)
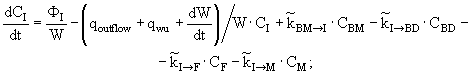 (13)
(13)
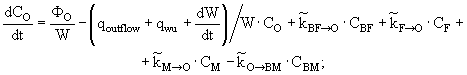 (14)
(14)
 (15)
(15)
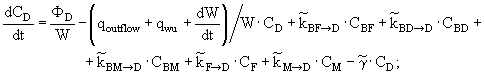 (16)
(16)
![]() . (17)
. (17)
Таким образом, определение динамики концентрации азота в пресноводных экосистемах сводится к решению задачи Коши для системы дифференциальных уравнений (10) - (17) с начальными условиями:
![]() , i = BF, BD, BM, I, O, F, D, M. (18)
, i = BF, BD, BM, I, O, F, D, M. (18)
Влияние температуры воды на исследуемые внутриводоемные процессы учитывается в виде зависимости констант (коэффициентов модели) от температуры. При описании круговорота азота в конкретных водоемах, обладающих специфическими особенностями, конкретный вид модели может существенно упрощаться.
В работе (Ross et al., 1994) рассматривается несколько вариантов экосистемных имитационных моделей морских заливов. Отличительной чертой данных моделей является учет эффектов самозатенения фитопланктона, процессов агрегирования зоопланктона в районах с относительно более высокой плотностью фитопланктонной биомассы, миграции зоопланктона и хищников в различные слои воды, более детальное описание жизненного цикла зоопланктона в зависимости от сезона и температуры воды, учет процессов перемешивания, апвеллинга, речного стока.
Цель работы состояла в том, чтобы получить количественное соответствие результатов модельных расчетов с имеющимися данными наблюдений для заливов, значительно отличающихся по своему гидрологическому и гидробиологическому режиму: средние глубины - от 8 до 150 м; различное время водообмена с морем - от одних до 10-14 суток; разное время наступления, продолжительность и интенсивность весеннего пика биомассы фитопланктона. При этом расчеты для всех водоемов выполнялись при одном и том же наборе параметров.
Предполагалось, что вертикальная структура воды состоит из трех слоев: верхний слой - от поверхности воды до верхней границы пикноклина, промежуточный слой - от верхней границы пикноклина до его нижней границы, придонный слой - от нижней границы пикноклина до дна. Поскольку детальная информация о сезонной изменчивости границ пикноклина отсутствует, при моделировании предполагалось, что все три слоя имеют одинаковую толщину. Кроме того, принималось допущение о том, что все три слоя вертикально и горизонтально однородны. Верхний и промежуточный слои связаны с морем в результате действия приливов. Кроме этого, каждый из этих слоев связан друг с другом процессами вертикального перемешивания и приливного апвеллинга. В верхний слой воды поступают воды с речным стоком и привносят в экосистему биогенные элементы. Процессы перемешивания, апвеллинга, водообмена с морем, речного стока описывались в модели реакциями первого порядка.
Биотические компоненты экосистемы были разделены на три функциональные группы: фитопланктон, зоопланктон и хищники. Фитопланктон поверхностного и промежуточного слоев рассматривался раздельно, но при этом учитывались процессы осаждения, апвеллинга и турбулентного перемешивания. Предполагалось также, что фитопланктон поглощает неорганический азот и выделяет органический азот только в пределах своего слоя. Считалось, что фитопланктон, который опустился ниже нижней границы пикноклина, отмирает и пополняет азотный пул придонного слоя. Зоопланктон питается фитопланктоном как в поверхностном, так и в промежуточном слоях и выделяет неорганический азот в оба эти слоя в количествах, пропорциональных времени, которое он проводит в каждом из них. Предполагалось, что трупы зоопланктона и фекальные пеллеты быстро опускаются в придонный слой воды и, таким образом, выделение азота происходит непосредственно в этом слое. Хищники питаются зоопланктоном. Все другие процессы для них описываются точно также, как и для зоопланктона. В модели считалось, что процессы первичного продуцирования лимитируются или световыми условиями или внутриклеточной квотой азота (N/C), в то время как образование вторичной продукции ограничивается только доступностью углерода.
Поскольку предполагалось, что поверхностный и промежуточный слои воды вертикально и горизонтально однородны, то распределение фитопланктона, концентрации биогенных элементов и азотной квоты также однородно в пределах каждого из этих слоев. Самозатенение фитопланктона учитывалось при расчете коэффициента поглощения. Показано, что если не учитывать этот эффект, то для периода весеннего цветения модельные оценки и данные реальных наблюдений могут различаться почти в два раза. Вторым механизмом, включенным в модель и регулирующим формирование нереалистично высоких концентраций фитопланктона, является агрегирование зоопланктона в зонах повышения концентрации фитопланктона ("top-down" control).
Процессы роста и репродукции зоопланктона тесно связаны с температурой воды и, следовательно, с сезоном года. Динамика этих процессов связана также с образованием покоящихся яиц. В течение зимы основное пополнение популяции зоопланктона происходит за счет яиц, покоящихся в донных отложениях (ДО). В остальное время года большинство яиц вылупляется непосредственно воде. Максимальное поступление покоящихся яиц в ДО имеет место в июле и августе. Таким образом, в модели учитывается два важнейших процесса: 1) изменение биомассы зоопланктона за счет процессов роста, прижизненных выделений, смертности и т. п.; 2) изменение численности и биомассы зоопланктона за счет образования покоящихся яиц.
Математическое описание модели динамики экосистемы фьордов.
Таблица 1. Переменные модели.
|
Обозначение |
Определение |
Единица измерения |
|
|
Биомасса фитопланктона в единицах углерода в поверхностном слое воды |
mgC m-3 |
|
|
Биомасса фитопланктона в единицах азота в поверхностном слое воды |
mgN m-3 |
|
|
Биомасса фитопланктона в единицах углерода в промежуточном слое воды |
mgC m-3 |
|
|
Биомасса фитопланктона в единицах азота в промежуточном слое воды |
mgN m-3 |
|
|
Биомасса зоопланктона в единицах углерода |
mgC m-3 |
|
|
Биомасса зоопланктона в единицах азота |
mgN m-3 |
|
|
Биомасса хищников в единицах углерода |
mgC m-3 |
|
|
Биомасса хищников в единицах азота |
mgN m-3 |
|
|
Концентрация DIN в поверхностном слое воды |
mgN m-3 |
|
|
Концентрация DIN в промежуточном слое воды |
mgN m-3 |
|
|
Концентрация DIN в придонном слое воды |
mgN m-3 |
|
|
Концентрация DON в поверхностном слое воды |
mgN m-3 |
|
|
Концентрация DON в промежуточном слое воды |
mgN m-3 |
Таблица 2. Параметры модели.
|
Обозначение |
Определение |
Величина |
Единица измерения |
|
|
Константы в функции, описывающей скорость образования яиц зоопланктона |
100; 100 |
d-1; °C-1 |
|
|
Максимальная скорость хищничества |
15 |
d-1 |
|
|
Фактор вымывания-удержания фитопланктона (Washout-Retention Factor) |
0.5 |
- |
|
|
Скорость смертности фитопланктона |
0.1 |
d-1 |
|
|
Скорость смертности зоопланктона |
0.05 |
d-1 |
|
|
Скорость смертности хищников |
0.05 |
d-1 |
|
|
Коэффициенты температурной функции |
1.0; 0.1 |
-; °C-1 |
|
|
Коэффициенты температурной функции |
1.0; 0.1 |
-; °C-1 |
|
|
Коэффициенты температурной функции |
1.0; 0.1 |
-; °C-1 |
|
|
Доля времени, которое хищник проводит в поверхностном слое воды |
0.5 |
d-1 |
|
|
Константы в функции, описывающей вертикальную миграцию зоопланктона |
0.5; 100 |
-; - |
|
|
Доля поглощенной пищи, теряемой зоопланктоном в результате дефекации |
0.36 |
- |
|
|
Доля поглощенной пищи, теряемой хищниками в результате дефекации |
0.5 |
- |
|
|
Доля биомассы фитопланктона, теряемой в результате обменных процессов |
0.25 |
d-1 |
|
|
Доля биомассы зоопланктона, теряемой в результате обменных процессов |
0.05 |
d-1 |
|
|
Доля биомассы хищников, теряемой в результате обменных процессов |
0.75 |
d-1 |
|
|
Доля поглощенной пищи, теряемой фитопланктоном в результате процессов выделения |
0.05 |
- |
|
|
Доля поглощенной пищи, теряемой зоопланктоном в результате процессов выделения |
0.15 |
- |
|
|
Доля поглощенной пищи, теряемой хищниками в результате процессов выделения |
0.2 |
- |
|
|
Максимальная скорость выедания зоопланктоном фитопланктона |
2.0 |
d-1 |
|
|
Коэффициент полунасыщения при поглощении DIP фитопланктоном |
4.2 |
mgN m-3 |
|
|
Коэффициент полунасыщения при поглощении света фитопланктоном |
60 |
Einst m-2 h-1 |
|
|
Коэффициент полунасыщения при поглощении фитопланктона зоопланктоном |
150 |
mgC m-3 |
|
|
Коэффициент полунасыщения при поглощении зоопланктона хищниками |
400 |
mgC m-3 |
|
|
Константа скорости реминерализации на границе раздела вода - ДО |
0.01 |
d-1 |
|
|
Константа скорости реминерализации DON |
0.02 |
d-1 |
|
|
Азотная квота для иммигрирующего фитопланктона |
0.15 |
mgN(mgC)-1 |
|
|
Азотная квота для иммигрирующего зоопланктона |
0.15 |
mgN(mgC)-1 |
|
|
Азотная квота для иммигрирующих хищников |
0.15 |
mgN(mgC)-1 |
|
|
Максимальная квота для фитопланктона по азоту |
0.25 |
mgN(mgC)-1 |
|
|
Минимальная квота для фитопланктона по азоту |
0.05 |
mgN(mgC)-1 |
|
|
Предельная квота для фитопланктона по азоту |
0.01 |
mgN(mgC)-1 |
|
|
Максимальная скорость поглощения азота фитопланктоном на единицу биомассы |
1.2 |
mgN(mgC)-1 d-1 |
|
|
Максимальная скорость роста фитопланктона при лимитировании азотом |
1.6 |
d-1 |
|
|
Максимальная скорость роста фитопланктона при лимитировании световыми условиями |
1.2 |
d-1 |
|
|
Вклад фитопланктона в коэффициент поглощения |
0.012 |
m2(mgC)-1 |
Таблица 3. Скорости процессов.
|
Обозначение |
Определение |
Единица измерения |
|
|
Общая скорость поглощения зоопланктона хищниками |
mgC d-1 |
|
|
Глубина в пределах поверхностного слоя воды, на которой свет становится лимитирующим фактором |
m |
|
|
Глубина в пределах промежуточного слоя воды, на которой свет становится лимитирующим фактором |
m |
|
|
Коэффициенты поглощения PAR в поверхностном и промежуточном слоях |
m-1 |
|
|
Доля времени, проводимая зоопланктоном в поверхностном слое воды |
- |
|
|
Общая скорость образования трупов и фекальных пеллет |
mgN d-1 |
|
|
Осредненное по глубине лимитирование роста фитопланктона азотом в пределах поверхностного и промежуточного слоев |
d-1 |
|
|
Доля от поглощенной пищи, ассимилируемой фитопланктоном, зоопланктоном и хищниками |
- |
|
|
Общая скорость поглощения зоопланктоном фитопланктона в пределах поверхностного и промежуточного слоев воды |
mgC d-1 |
|
|
Суточные потери биомассы фито-, зоопланктона и хищников в результате смертности, процессов прижизненных выделений и репродукции |
d-1 |
|
|
Азотные квоты для фитопланктона в поверхностном и промежуточном слоях воды |
mgN (mgC)-1 |
|
|
Азотные квоты для зоопланктона в поверхностном и промежуточном слоях воды |
mgN (mgC)-1 |
|
|
Температурные функции для фито-, зоопланктона и хищников |
- |
|
|
Общая скорость поглощения углерода фитопланктоном в поверхностном и промежуточном слоях воды |
mgC d-1 |
|
|
Общая скорость поглощения азота фитопланктоном в поверхностном и промежуточном слоях воды |
mgN d-1 |
|
|
Общая скорость прижизненных выделений фитопланктона в поверхностном и промежуточном слоях воды |
mgN d-1 |
|
|
Общая скорость прижизненных выделений зоопланктона в поверхностном и промежуточном слоях воды |
mgN d-1 |
|
|
Общая скорость прижизненных выделений хищников в поверхностном и промежуточном слоях воды |
mgN d-1 |
Таблица 4. Физические параметры.
|
Обозначение |
Определение |
Единица измерения |
|
|
Объемы поверхностного, промежуточного и придонного слоев воды |
m3 |
|
|
Доля приливного апвеллинга в пределах поверхностного слоя воды |
- |
|
|
Скорость водообмена в поверхностном слое воды в результате действия приливов |
m3 d-1 |
|
|
Скорость водообмена в поверхностном слое воды в результате стока поверхностных вод |
m3 d-1 |
|
|
Скорость водообмена в промежуточном слое воды в результате стока поверхностных вод |
m3 d-1 |
|
|
Коэффициент поглощения света водой |
m-1 |
|
|
Глубина поверхностного и промежуточного слоев воды |
m |
|
|
Солнечная радиация у поверхности воды |
Einst m-2 h-1 |
|
|
Температура воды |
°С |
Уравнения модели.
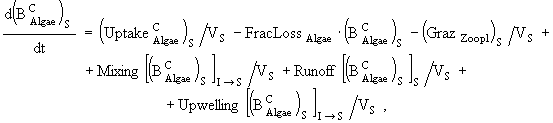 (1)
(1)
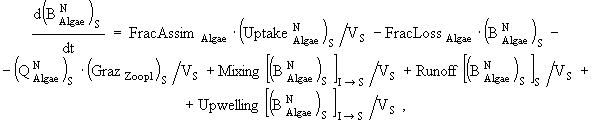 (2)
(2)
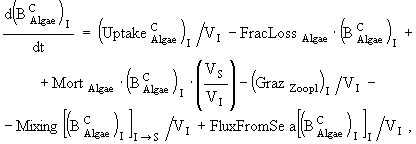 (3)
(3)
 (4)
(4)
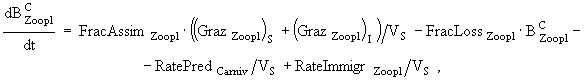 (5)
(5)
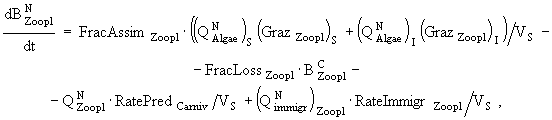 (6)
(6)
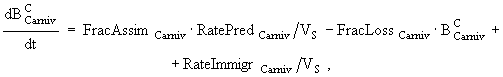 (7)
(7)
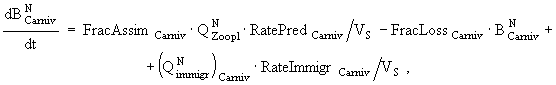 (8)
(8)
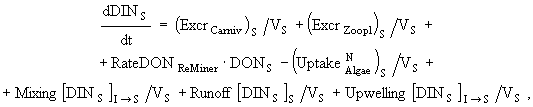 (9)
(9)
 (10)
(10)
![]() (11)
(11)
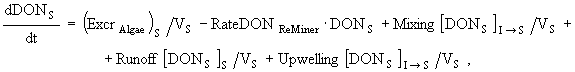 (12)
(12)
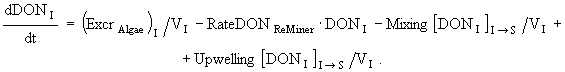 (13)
(13)
Параметризация скоростей процессов.
![]() (14)
(14)
![]()
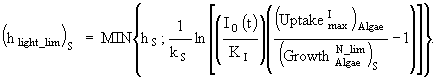 (15)
(15)
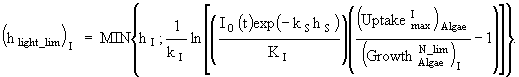 (16)
(16)
![]() (17)
(17)
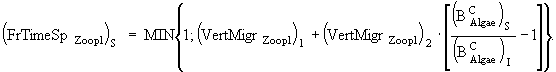 (18)
(18)
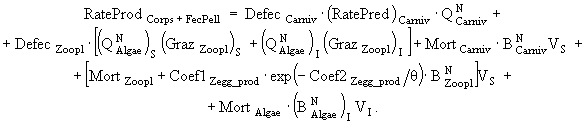 (19)
(19)
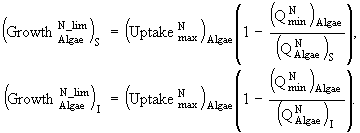 (20)
(20)
 (21)
(21)
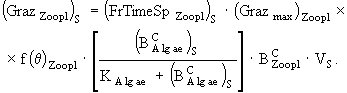 (22)
(22)
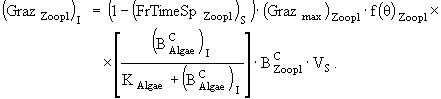 (23)
(23)
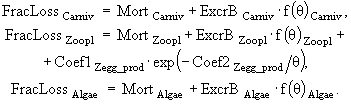 (24)
(24)
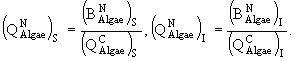 (25)
(25)
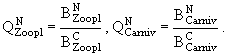 (26)
(26)
 (27)
(27)
![]()
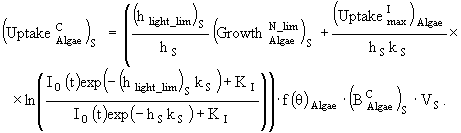 (28)
(28)
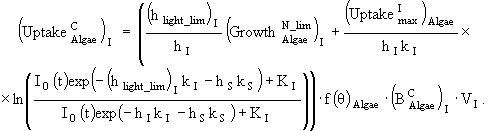 (29)
(29)
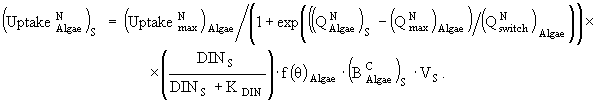 (30)
(30)
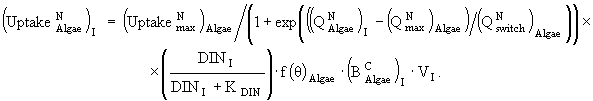 (31)
(31)
 (32)
(32)
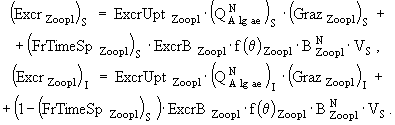 (33)
(33)
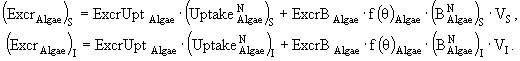 (34)
(34)
·С помощью трехмерной математической модели в работе (Fennel, 1999) исследовалось влияние вертикальной устойчивости столба жидкости на распространение весеннего цветения фитопланктона в западной части Балтийского моря. Модель включала в себя гидродинамический блок для расчета мезомасштабной циркуляции и простую биохимическую модель. Трехмерная модель циркуляции позволяет описывать конвективные процессы, поскольку в ней в явном виде учитывается зависимость плотности морской воды от температуры и солености. Известно, что начало периода цветения в Балтийском море с запада на восток сдвинуто во времени, хотя уровень приходящей солнечной радиации и содержание биогенных элементов отличаются в этих районах незначительно. Цель моделирования состояла в том, чтобы выяснить причины и механизм возникновения указанного временного сдвига. Пространственно-временная изменчивость исследовалась на модели в сравнении с данными наблюдений с 1994 по 1996 годы. Моделирование показало, что цветение начинается сразу после того, как закончится период весеннего конвективного перемешивания. Весенняя конвекция представляет собой обычное явление в солоноватых водах. Это происходит вследствие нелинейной зависимости плотности от температуры, но имеет место только тогда, когда температура воды станет ниже температуры максимальной плотности. Температура максимальной плотности более высокая в менее соленых водах восточной части Балтийского моря, поскольку критическая температура увеличивается с уменьшением солености. Таким образом, из-за градиента солености в восточном направлении и период конвективного перемешивания увеличивается в Балтийском море с запада на восток. Более продолжительный период перемешивания в восточных частях объясняет наблюдаемый западно-восточный временной лаг инициализации весеннего цветения фитопланктона. Внутригодовые различия в начале и продолжительности весеннего цветения могут в значительной степени влиять на трофодинамические процессы между фито- и зоопланктоном. Биохимическая модель включена в модель циркуляции посредством использования уравнений адвекции-диффузии для всех переменных состояния, которые рассматриваются как неконсервативные трассеры. В качестве лимитирующего биогенного элемента выбран азот. Скорость роста фитопланктона определяется модифицированной функцией Михаэлиса-Ментен, зоопланктона - модифицированной функцией Ивлева.
Уравнения химико-биологического блока модели:
·скорость изменения концентрации биогенного элемента (азота) -
![]() ; (1)
; (1)
·скорость изменения концентрации фитопланктона -
![]() ; (2)
; (2)
·скорость изменения концентрации зоопланктона -
![]() ; (4)
; (4)
·скорость изменения концентрации детрита -
![]() . (5)
. (5)
Здесь ![]() - скорость течения; А - коэффициент горизонтальной турбулентной диффузии;
- скорость течения; А - коэффициент горизонтальной турбулентной диффузии; ![]() - трехмерный оператор Лапласа; P0, Z0 - фоновые значения концентраций фито- и зоопланктона;
- трехмерный оператор Лапласа; P0, Z0 - фоновые значения концентраций фито- и зоопланктона; ![]() - максимальная скорость роста фитопланктона с коррекцией на температуру Т и условия освещенности I;
- максимальная скорость роста фитопланктона с коррекцией на температуру Т и условия освещенности I; ![]() - величина оптимальной освещенности;
- величина оптимальной освещенности; ![]() - скорость роста фитопланктона при температуре 0°С (0,8); a = 0,063 - эмпирический коэффициент;
- скорость роста фитопланктона при температуре 0°С (0,8); a = 0,063 - эмпирический коэффициент; ![]() - константа полунасыщения (0,09);
- константа полунасыщения (0,09); ![]() - максимальная скорость выедания зоопланктоном фитопланктона (0,5 сутки-1);
- максимальная скорость выедания зоопланктоном фитопланктона (0,5 сутки-1); ![]() - коэффициент Ивлева (1,1);
- коэффициент Ивлева (1,1); ![]() - скорость метаболических выделений фитопланктона с коррекцией на температуру;
- скорость метаболических выделений фитопланктона с коррекцией на температуру; ![]() - скорость выделения при температуре 0°С (0,06 сутки-1);
- скорость выделения при температуре 0°С (0,06 сутки-1); ![]() - скорость отмирания фитопланктона (0,02 сутки-1);
- скорость отмирания фитопланктона (0,02 сутки-1); ![]() ;
; ![]() - скорость дыхания зоопланктона (0,01 сутки-1);
- скорость дыхания зоопланктона (0,01 сутки-1); ![]() - скорость отмирания зоопланктона (0,02 сутки-1);
- скорость отмирания зоопланктона (0,02 сутки-1); ![]() ,
, ![]() - скорость реминерализации с коррекцией на температуру;
- скорость реминерализации с коррекцией на температуру; ![]() - скорость реминерализации при температуре воды 0°С (0,1 сутки-1);
- скорость реминерализации при температуре воды 0°С (0,1 сутки-1); ![]() - скорость оседания фитопланктона; b = 120 см / сутки;
- скорость оседания фитопланктона; b = 120 см / сутки; ![]() - скорость оседания детрита (500 см / сутки).
- скорость оседания детрита (500 см / сутки).
·В работе (Gin et al., 1998) предлагается теоретическая модель, рассматривающая размерную структуру планктонных сообществ, особенности их функционирования, переноса энергии (биомассы) между разными трофическими уровнями и реакции на различные возмущения факторов среды. Модель используется для моделирования изменений размерной структуры фитопланктона и первичной продукции в соответствии с изменениями вертикальной структуры воды.
Азот в данной модели рассматривается в качестве единственного лимитирующего биогенного элемента. Все моделируемые компартменты также рассматриваются в единицах азота. В модель включено шесть переменных состояния: 1) фитопланктон (Phy), 2) зоопланктон (Zoo), 3) азот нитратов (N), 4) аммонийный азот (A), 5) взвешенный органический азот (PON) и 6) растворенный органический азот (DON). Основные потоки включают в себя поглощение азота фитопланктоном, прижизненные метаболические выделения, хищничество, экскрецию, смертность и разложение взвешенного органического вещества. Поскольку бактериопланктон не рассматривается в модели явно, то его влияние моделируется опосредованно через цикл органического вещества и его разложение. Скорости изменения биомассы каждого из компартментов модели определяются скоростями поступления и выноса азота. На скорости процессов влияют внешние факторы, такие как внутрисуточные изменения световых условий и сезонные изменения вертикальной структуры воды. Фитопланктон и зоопланктон разделены в модели на семь размерных классов в логарифмическом масштабе (по основанию 2). Общий диапазон размерного изменения фитопланктона - 0,4 - 25,6 мкм, зоопланктона - 4 - 256 мкм ESD (Equivalent Spherical Diameter). Введение разных размерных классов дает возможность дифференцировать значения скоростей роста, оседания и питания.
Динамика каждого из компартментов представлена в модели балансовыми уравнениями
·Скорость изменения концентрации нитратного азота (N) -
![]() = -Photosynthesis?
= -Photosynthesis?![]() + Nitrification + Diffusion. (1)
+ Nitrification + Diffusion. (1)
Здесь ![]() - отношение поглощения нитратов к общему азоту (нитраты + аммонийный азот). Этот множитель задается далее в уравнении (13).
- отношение поглощения нитратов к общему азоту (нитраты + аммонийный азот). Этот множитель задается далее в уравнении (13).
·Скорость изменения концентрации аммонийного азота (A) -
![]() = - Photosynthesis?(1 -
= - Photosynthesis?(1 - ![]() ) - Nitrification + Decomposition of PON into Ammonium +
) - Nitrification + Decomposition of PON into Ammonium +
+ Decomposition of DON into Ammonium + Excretion of Zooplankton + Diffusion; (2)
·Скорость изменения концентрации взвешенного органического азота (PON) -
![]() = Mortality of Phytoplankton + Mortality of Zooplankton + Egestion of Zooplankton -Sinking of PON - Decomposition of PON into DON -
= Mortality of Phytoplankton + Mortality of Zooplankton + Egestion of Zooplankton -Sinking of PON - Decomposition of PON into DON -
- Decomposition of PON into Ammonium + Diffusion. (3)
·Скорость изменения концентрации растворенного органического азота (DON) -
![]() = Extracellular excretion of Phytoplankton + Decomposition of PON into DON -
= Extracellular excretion of Phytoplankton + Decomposition of PON into DON -
- Decomposition of DON into Ammonium + Diffusion. (4)
·Скорость изменения концентрации фитопланктона (Phy) -
![]() = Photosynthesis - Extracellular excretion - Mortality of Phytoplankton - Grazing -
= Photosynthesis - Extracellular excretion - Mortality of Phytoplankton - Grazing -
Sinking of Phytoplankton + Diffusion. (5)
·Скорость изменения концентрации зоопланктона (Zoo) -
![]() = Grazing - Egestion - Excretion - Mortality of Zooplankton + Diffusion. (6)
= Grazing - Egestion - Excretion - Mortality of Zooplankton + Diffusion. (6)
Рассмотрим теперь описание отдельных процессов. Скорость фотосинтеза определяется следующим соотношением:
Photosynthesis = ![]() , (7)
, (7)
где ![]() - удельная скорость фотосинтеза,
- удельная скорость фотосинтеза, ![]() - температурный коэффициент для коррекции скорости на температурные условия. Скорость фотосинтеза задается в соответствии с законом Либиха, то есть предполагается, что только один из лимитирующих факторов может ограничивать рост в данный момент времени:
- температурный коэффициент для коррекции скорости на температурные условия. Скорость фотосинтеза задается в соответствии с законом Либиха, то есть предполагается, что только один из лимитирующих факторов может ограничивать рост в данный момент времени:
![]() . (8)
. (8)
![]() является функций фотосинтетически активной радиации (PAR), которая, в свою очередь, зависит от времени года, широты места, глубины, способности воды поглощать свет, самозатенения фитопланктона и конкретного вида зависимости "Освещенность - Фотосинтез". В данной работе была использована функция Смита:
является функций фотосинтетически активной радиации (PAR), которая, в свою очередь, зависит от времени года, широты места, глубины, способности воды поглощать свет, самозатенения фитопланктона и конкретного вида зависимости "Освещенность - Фотосинтез". В данной работе была использована функция Смита:
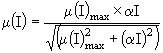 , (9)
, (9)
где ![]() - максимальное значение скорости роста при световом лимитировании, I - освещенность на заданной глубине,
- максимальное значение скорости роста при световом лимитировании, I - освещенность на заданной глубине, ![]() - начальный наклон P-Iкривой. Суточный ход освещенности на заданной глубине задавался уравнением:
- начальный наклон P-Iкривой. Суточный ход освещенности на заданной глубине задавался уравнением:
![]() . (10)
. (10)
Здесь ![]() - продолжительность дня, t - момент времени от нуля часов, k(Z) - коэффициент поглощения света водой и фитопланктоном в результате процесса самозатенения:
- продолжительность дня, t - момент времени от нуля часов, k(Z) - коэффициент поглощения света водой и фитопланктоном в результате процесса самозатенения:
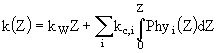 , (11)
, (11)
где ![]() - коэффициент поглощения света фитопланктоном разных размерных классов. Функция
- коэффициент поглощения света фитопланктоном разных размерных классов. Функция ![]() вычисляется по формуле
вычисляется по формуле
 , (12)
, (12)
то есть для описания скорости роста, лимитированного питательными веществами, было использовано уравнение Михаэлиса-Ментен. Поскольку у фитопланктона существует пищевое предпочтение аммиачного азота то, следовательно, будет иметь место ингибирование поглощения нитратов в присутствии значительных концентраций ![]() . Это обстоятельство и учтено в уравнении (12) путем введения экспоненциального множителя в первом слагаемом (
. Это обстоятельство и учтено в уравнении (12) путем введения экспоненциального множителя в первом слагаемом (![]() - фактор ингибирования; в данной модели принято, что
- фактор ингибирования; в данной модели принято, что ![]() =1,5). Отношение поглощения нитратов к общему азоту задается следующим соотношением:
=1,5). Отношение поглощения нитратов к общему азоту задается следующим соотношением:
![]() . (13)
. (13)
Внеклеточные выделения представляют собой долю чистой первичной продукции, выделяемой фитопланктоном в виде DON. В модели предполагается, что
Extracellular excretion = ![]() ?Photosynthesis, (14)
?Photosynthesis, (14)
где ![]() - удельная скорость выделения. Предполагается, что процесс выедания зоопланктоном клеток фитопланктона представляет собой размерно-зависимый процесс, в котором каждый размерный класс зоопланктона выедает только тот размерный класс фитопланктона, который является ближайшим к данному классу. Скорость выедания задается в модели следующей гиперболической функцией:
- удельная скорость выделения. Предполагается, что процесс выедания зоопланктоном клеток фитопланктона представляет собой размерно-зависимый процесс, в котором каждый размерный класс зоопланктона выедает только тот размерный класс фитопланктона, который является ближайшим к данному классу. Скорость выедания задается в модели следующей гиперболической функцией:
![]() . (15)
. (15)
Здесь ![]() - максимальное значение удельной скорости выедания, слагаемое refugeопределяет пороговое значение, ниже которого выедание данного размерного класса фитопланктона прекращается. Метаболические потери углерода в результате дыхания согласовываются в модели через эквивалентные потери азота так, чтобы оставалось постоянным соотношение C:Nдля гетеротрофов. В модели принято, что выделение пропорционально скорости выедания:
- максимальное значение удельной скорости выедания, слагаемое refugeопределяет пороговое значение, ниже которого выедание данного размерного класса фитопланктона прекращается. Метаболические потери углерода в результате дыхания согласовываются в модели через эквивалентные потери азота так, чтобы оставалось постоянным соотношение C:Nдля гетеротрофов. В модели принято, что выделение пропорционально скорости выедания:
Excretion = ![]() ?Grazing, (16)
?Grazing, (16)
где ![]() - удельная скорость выделения. Доля съеденного материала, который гетеротрофы не усвоили, выделяется в виде фекалиев:
- удельная скорость выделения. Доля съеденного материала, который гетеротрофы не усвоили, выделяется в виде фекалиев:
Egestion = (1 - ![]() )Grazing, (17)
)Grazing, (17)
где ![]() - коэффициент ассимиляции пищи гетеротрофами. Этот коэффициент одинаков для всех размерных классов зоопланктона. Рециклинг питательных веществ в модели возникает в результате процесса отмирания фитопланктона и зоопланктона. Предполагается, что этот процесс зависит от температуры и пропорционален второй степени концентрации планктона:
- коэффициент ассимиляции пищи гетеротрофами. Этот коэффициент одинаков для всех размерных классов зоопланктона. Рециклинг питательных веществ в модели возникает в результате процесса отмирания фитопланктона и зоопланктона. Предполагается, что этот процесс зависит от температуры и пропорционален второй степени концентрации планктона:
Mortality of Phytoplankton = ![]() , (18)
, (18)
Mortality of Zooplankton = ![]() , (19)
, (19)
где ![]() - удельные скорости смертности фито- и зоопланктона при нуле градусов. Смертность планктона формирует пулы органического углерода (DONи PON), которые в результате их минерализации бактериями превращаются в аммонийный азот. Чтобы не усложнять модель и не вводить бактериальный компартмент, для описания разложения форм органического азота используется линейная зависимость с коррекцией на температурные условия:
- удельные скорости смертности фито- и зоопланктона при нуле градусов. Смертность планктона формирует пулы органического углерода (DONи PON), которые в результате их минерализации бактериями превращаются в аммонийный азот. Чтобы не усложнять модель и не вводить бактериальный компартмент, для описания разложения форм органического азота используется линейная зависимость с коррекцией на температурные условия:
Decomposition of PON into Ammonium = ![]() , (20)
, (20)
Decomposition of PON into DON = ![]() , (21)
, (21)
Decomposition of DON into Ammonium = ![]() . (22)
. (22)
Аммиачный азот далее превращается в нитратный азот в присутствии кислорода (процесс нитрификации):
Nitrification = ![]() . (23)
. (23)
Для описания процесса оседания фитопланктона использовалось аллометрическое соотношение между скоростью оседания ![]() и массой клеток, полученное по эмпирическим данным при исследовании морского фитопланктона. Масса клеток Wвыражалась в единицах углерода и менялась от 0 до 30000 пикограмм С:
и массой клеток, полученное по эмпирическим данным при исследовании морского фитопланктона. Масса клеток Wвыражалась в единицах углерода и менялась от 0 до 30000 пикограмм С:
![]() (м / сутки). (24)
(м / сутки). (24)
Таким образом, оседание фитопланктона задается выражением:
![]() . (25)
. (25)
Оседание взвешенного органического азота задается аналогично:
![]() , (26)
, (26)
причем ![]() = 1 м / сутки в пределах верхнего перемешанного слоя и
= 1 м / сутки в пределах верхнего перемешанного слоя и ![]() = 10 м / сутки ниже этого слоя.
= 10 м / сутки ниже этого слоя.
Размер фитопланктона и зоопланктона выражался либо в объемных единицах, либо в единицах ESD. Для расчета его биомассы, объемный размер переводился в единицы углерода, для чего использовались различные эмпирические зависимости. Скорости метаболических процессов, такие как удельная скорость поглощения питательных веществ и скорость питания, рассматриваются в модели как функция текущего размерного класса i:
![]() , (27)
, (27)
где символ 0 относится к самому маленькому размерному классу, а биомасса организмов выражается в единицах углерода. ![]() - эмпирическая постоянная, значение которой принималось равным -0,25. Константы полунасыщения также зависят от размерного класса. При описании поглощения биогенных элементов фитопланктоном, использовалось одно и то же значение
- эмпирическая постоянная, значение которой принималось равным -0,25. Константы полунасыщения также зависят от размерного класса. При описании поглощения биогенных элементов фитопланктоном, использовалось одно и то же значение ![]() и для потребления нитратного и для потребления аммонийного азота:
и для потребления нитратного и для потребления аммонийного азота:
![]() . (28)
. (28)
В случае зоопланктона предполагалось, что константа полунасыщения не зависит от его размера, но зависит от размерного класса фитопланктона:
![]() . (29)
. (29)
![]() , вычисленная с помощью соотношения (29), переводилась затем в единицы азота с помощью отношения Редфилда 6,625.
, вычисленная с помощью соотношения (29), переводилась затем в единицы азота с помощью отношения Редфилда 6,625.
При создании математической модели круговорота фосфора в пресноводных экосистемах в работе (Добрынский, Рогаль, 1993) выделяли следующие укрупненные компоненты экосистемы: 1) водные растворы минеральных неорганических соединений фосфора; 2) водорастворимые органические формы фосфора; 3) взвеси водонерастворимых веществ типа детрита и твердые органические остатки; 4) фитопланктон; 5) зоопланктон; 6) бактериопланктон; 7) перифитон; 8) бентос; 9) высшая водная растительность (макрофиты); 10) ихтиофауна. Происходящие в пресноводных экосистемах циклы внутриводоемной биохимической трансформации соединений фосфора являются следствием одновременно протекающих процессов продукции и деструкции органического вещества. При этом происходит как потребление биотой соединений фосфора, так и выделение продуктов метаболизма и образование (за счет отмирания живых организмов) детрита. При построении данной математической модели биомасса зоопланктона и ихтиофауны, а также их влияние на внутриводоемные процессы трансформации вещества не учитываются. Принимая во внимание относительное постоянство биомассы макрофитов, их участие во внутриводоемных процессах в модели представлено как действие некоторого, вполне определенного по величине внешнего фактора. Полагается также, что каждая компонента фосфорной системы характеризуется своей концентрацией. При этом под концентрацией таких компонент, как фосфор фитопланктона, бактериальный, детрита и макрофитов, подразумевается отношение общего количества в сырой массе вещества каждой из указанных компонент к тому объему воды, в котором они рассматриваются. Основные гипотезы (предположения), на основании которых производится вывод уравнений, описывающих круговорот фосфора в пресноводных экосистемах, состоят в следующем:
1. Неорганический фосфор поглощается макрофитами в количестве, пропорциональном биомассе макрофитов и концентрации минеральных форм фосфора.
2. Водорастворимые органические формы фосфора поглощаются бактериями. При этом выделяется неорганический фосфор, то есть происходит трансформация органического фосфора в неорганический. Количество выделившегося неорганического фосфора пропорционально биомассе бактерий и концентрации органического фосфора.
3. Фитопланктон, поглощая неорганический фосфор, выделяет водорастворимые формы органического фосфора в количестве, пропорциональном биомассе фитопланктона и концентрации неорганического фосфора.
4. В процессе разложения мертвого органического вещества (детрита) образуется органический фосфор. Его количество пропорционально массе детрита.
5. В процессе своей жизнедеятельности высшая водная растительность выделяет водорастворимые органические соединения фосфора в количестве, пропорциональном биомассе макрофитов.
6. Смертность фитопланктона пропорциональна его биомассе. Коэффициент пропорциональности (смертности) зависит от отношения показателей концентрации фитопланктона и неорганического фосфора. При этом увеличивается масса детрита и уменьшается биомасса фитопланктона.
7. Смертность бактерий пропорциональна их биомассе. Коэффициент пропорциональности зависит от отношения концентрации бактерий и концентрации водорастворимых органических соединений фосфора. При этом увеличивается масса детрита и уменьшается биомасса бактерий.
8. Часть детритных соединений фосфора в процессе седиментации депонируется в донных отложениях.
9. Компоненты фосфорной системы равномерно распределены по объему водоема (водотока), то есть их концентрация не зависит от пространственных координат.
Влияние температуры воды на исследуемые водоемные процессы продукции и деструкции вещества учитывается через зависимости констант (или коэффициентов) от температуры.
В работе (Menshutkin et al., 1998) представлена новая версия экосистемной модели Ладожского озера. Модель относится только к пелагической части озера. Таким образом, влияние бентоса и высшей водной растительности не учитывается. Модель построена таким образом, чтобы воспроизводить только те воздействия и масштабы природных явлений, которые фактически наблюдаются при использовании современных лимнологических методов исследования. В соответствии с этим положением, не учитываются внутрисуточные изменения биотических и абиотических компонентов, локальные пространственные неоднородности концентрация фито- и зоопланктона, не рассматривается процесс внутрисуточных вертикальных миграций зоопланктона.
В модели рассматриваются три наиболее важные группы фитопланктона. Это представляет собой некоторый компромисс между одной группой в моделях Астраханцева с соавторами (Astrakhantsev et al., 1996), Меншуткина и Воробьевой (Меншуткин, Воробьева, 1987) и восемью группами фитопланктона, как это было в модели Воробьевой и Меншуткина (Воробьева, Меншуткин, 1989). Такая структура была выбрана с тем, чтобы на основе существующих данных наблюдений и специфики функционирования отдельных групп водных растений показать роль сукцессии фитопланктона в трансформации экосистемы. Первая группа состоит из видов, которые превалируют в фитопланктонном сообществе в весенний период (весенний диатомовый комплекс - Aulacosiza-Diatoma, Asterionella и другие) Вторая группа включает летние виды, где доминируют сине-зеленые водоросли (Microcystis, Oscillatoria). Третью группу формируют виды осеннего фитопланктонного комплекса (Woronichinia и Aulacosiza, Tribonema, Aphanizomenon). Зоопланктон в модели представлен как единая система, которая характеризуется значением общей биомассы. Это было вызвано необходимостью получить по крайней мере приблизительный баланс в точности отдельных частей модели. По этим же соображениям представленная модель не включает в себя подмодель рыбного сообщества, хотя и имеется хорошо разработанная модель ихтиоценоза Ладожского озера (Ладанов, Тихонов, 1985). Абиотическая часть модели не включает в себя рассмотрение круговорота азота, а рассматривается только фосфорный баланс, поскольку во многих работах было показано, что основным лимитирующим элементом в Ладожском озере является именно фосфор. Такой подход позволил уменьшить число переменных в модели. Кроме этого переменными модели являются растворенное ОВ, фосфор в составе детрита, растворенный кислород. Новая версия модели позволяет повысить пространственное разрешение по вертикали, поскольку предоставляет возможность работать с многослойной моделью. Учитываются также процессы переноса компонентов экосистемы течениями, турбулентного перемешивания и седиментации.
Краткое математическое описание модели экосистемы Ладожского озера.
Таблица 1. Переменные модели.
|
Обозначение |
Определение |
Единица измерения |
|
|
Биомасса трех видов водорослей |
mg l-1 |
|
|
Биомасса зоопланктона |
mg l-1 |
|
|
Концентрация фосфора в составе детрита |
mg l-1 |
|
|
Концентрация растворенного органического фосфора |
mg l-1 |
|
|
Концентрация растворенного ОВ |
mg l-1 |
|
|
Концентрация растворенного в воде кислорода |
mg l-1 |
Таблица 2. Скорости процессов.
|
Обозначение |
Определение |
Единица измерения |
|
|
Изменение биомассы фитопланктона за счет адвективного переноса |
mg l-1 day-1 |
|
|
Изменение биомассы зоопланктона за счет адвективного переноса |
mg l-1 day-1 |
|
|
Изменение концентрации фосфора в составе детрита за счет адвективного переноса |
mg l-1 day-1 |
|
|
Изменение концентрации растворенного органического фосфора за счет адвективного переноса |
mg l-1 day-1 |
|
|
Изменение концентрации растворенного органического вещества за счет адвективного переноса |
mg l-1 day-1 |
|
|
Изменение концентрации растворенного в воде кислорода за счет адвективного переноса |
mg l-1 day-1 |
|
|
Изменение биомассы фитопланктона за счет диффузионного переноса |
mg l-1 day-1 |
|
|
Изменение биомассы зоопланктона за счет диффузионного переноса |
mg l-1 day-1 |
|
|
Изменение концентрации фосфора в составе детрита за счет диффузионного переноса |
mg l-1 day-1 |
|
|
Изменение концентрации растворенного органического фосфора за счет дифузионного переноса |
mg l-1 day-1 |
|
|
Изменение концентрации растворенного органического вещества за счет диффузионного переноса |
mg l-1 day-1 |
|
|
Изменение концентрации растворенного в воде кислорода за счет диффузионного переноса |
mg l-1 day-1 |
|
|
Скорость образования первичной продукции фитопланктона |
mg l-1 day-1 |
|
|
Скорость смертности фитопланктона |
mg l-1 day-1 |
|
|
Скорость выедания фитопланктона зоопланктоном |
mg l-1 day-1 |
|
|
Скорость роста зоопланктона |
mg l-1 day-1 |
|
|
Скорость метаболических выделений зоопланктона |
mg l-1 day-1 |
|
|
Скорость смертности зоопланктона |
mg l-1 day-1 |
|
|
Скорость трансформации фосфора в составе детрита в растворенный неорганический фосфор |
mg l-1 day-1 |
|
|
Скорость трансформации фосфора в составе детрита в растворенный органический фосфор |
mg l-1 day-1 |
|
|
Скорость выедания детрита зоопланктоном |
mg l-1 day-1 |
|
|
Скорость разложения DOP |
mg l-1 day-1 |
Уравнения модели.
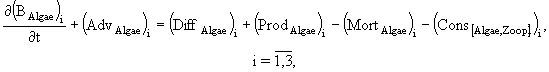 (1)
(1)
![]() (2)
(2)
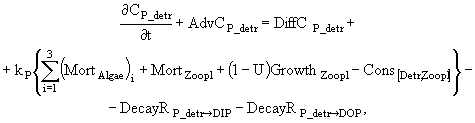 (3)
(3)
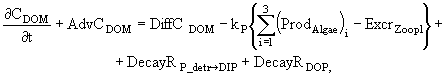 (4)
(4)
![]() (5)
(5)
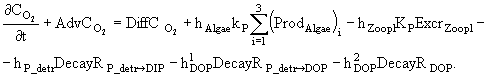 (6)
(6)
Параметризация скоростей отдельных процессов.
Адвективный перенос модельных компонентов экосистемы задается следующим выражением:
![]() (7)
(7)
где ![]() - трехмерный вектор скоростей течений,
- трехмерный вектор скоростей течений, ![]() - скорость седиментации, причем
- скорость седиментации, причем ![]() . Диффузионный перенос представлен следующим соотношением:
. Диффузионный перенос представлен следующим соотношением:
![]() (8)
(8)
где ![]() - коэффициенты турбулентной диффузии.
- коэффициенты турбулентной диффузии.
Расчет концентрации растворенного в воде кислорода при насыщении в зависимости от температуры воды осуществляется по формуле:
![]() (9)
(9)
Граничные условия.
1) Свободная поверхность -
![]() (10)
(10)
где ![]() - заданный поток компонента С через поверхность. Для кислорода это граничное условие записывается в виде
- заданный поток компонента С через поверхность. Для кислорода это граничное условие записывается в виде ![]() где
где ![]() - коэффициент аэрации,
- коэффициент аэрации, ![]() концентрация растворенного в воде кислорода при насыщении.
концентрация растворенного в воде кислорода при насыщении.
2) Дно водоема и твердая боковая граница -
![]() (11)
(11)
Для кислорода граничное условие задается в виде соотношения ![]()
3) Место впадения реки. Для С = {фосфор в составе детрита; DOP; DOM и кислорода}
![]() (12)
(12)
Для остальных компонентов системы -
![]() (13)
(13)
Таблица 3. Отдельные параметры модели.
|
Обозначение |
Определение |
Величина |
Единица измерения |
|
|
Фитопланктон первой группы |
|
|
|
|
Минимальная температура воды |
0.2 |
°C |
|
|
Максимальная температура воды |
6.0 |
°C |
|
|
Фитопланктон второй группы |
|
|
|
|
Минимальная температура воды |
0.1 |
°C |
|
|
Максимальная температура воды |
20.0 |
°C |
|
|
Фитопланктон третьей группы |
|
|
|
|
Минимальная температура воды |
10.0 |
°C |
|
|
Максимальная температура воды |
16.0 |
°C |
|
|
Коэффициент смертности фитопланктона (минимальный) |
0.05 |
day-1 |
|
|
Коэффициент смертности фитопланктона (максимальный) |
0.2 |
day-1 |
|
U |
Коэффициент усвоения пищи зоопланктоном |
0.75 |
- |
|
|
Коэффициент смертности зоопланктона (минимальный) |
0.05 |
day-1 |
|
|
Коэффициент смертности зоопланктона (максимальный) |
0.3 |
day-1 |
В работе (Савчук и др., 1982) построена резервуарная модель круговорота вещества в пелагической экосистеме Северного моря. Экосистема считается горизонтально однородной, а ее вертикальная структура аппроксимирована двумя гомогенными слоями, различающимися между собой концентрациями и условиями функционирования компонентов экосистемы. При описании планктонного сообщества компонентами модели служат экологически однородные группы фитопланктона и зоопланктона. Рассматриваемые как элементы единой системы, такие группы должны быть внутренне однородными в отношении связей со средой и различаться между собой характером и интенсивностью взаимодействия с другими элементами системы. При сегментации реального многовидового сообщества на ограниченное число экологически однородных групп важно, чтобы различия экологических характеристик между группами были бы существенно большими, чем экологические различия среди объединенных в одну группу видов. Выделение групп проведено на основе анализа структуры планктонного сообщества Северного моря. На основании таких критериев как объем клеток, фенологическое сходство отдельных видов, специфика потребления их животными и, в меньшей степени, систематическая принадлежность, выделены три экологически однородные группы фитопланктона: мелкоклеточный фитопланктон, крупные диатомовые и панцирные перидиниевые. По совокупности весовых, трофических и систематических характеристик выделены пять групп зоопланктона: тонкие фильтраторы, младшие копеподы весенних форм, старшие копеподы весенних форм, фитофаги летнего комплекса, хищный зоопланктон. В органическом веществе выделены взвешенный и растворенный компоненты, различающиеся своим отношением к факторам механического переноса (седиментация и перемешивание). При этом допускается, что детрит может служить источником пищи для зоопланктона, а растворенное органическое вещество (РОВ) - нет. Для модельного отражения природной биохимической разнородности детрита и РОВ каждый из этих компонентов подразделен на лабильную и стойкую фракции. Минеральные соединения представлены фосфатами.
При моделировании рассматриваются следующие процессы:
рост, естественная смертность, выедание зоопланктоном и гравитационное оседание клеток фитопланктона;
рост, естественная смертность, выедание более мелких групп зоопланктона более крупными группами;
экскреция фосфора зоопланктерами;
детритообразование в результате естественной смертности фито- и зоопланктона и поступления остатков пищевых частиц, "отброшенных" зоопланктоном;
деструкции и минерализации веществ, моделируемых как реакции первого порядка в зависимости от температуры воды;
обмен компонентами между слоями;
онтогенетический метаморфоз, который интерпретируется в модели как постоянный переход определенной доли биомассы "младшей" группы зоопланктона в биомассу "старшей".
Интересной особенностью данной модели является введение обратной связи между скоростью роста и численностью клеток фитопланктона. Этот подход означает непосредственный - в отличие от косвенного, через концентрацию биогенов - учет явлений "внутригрупповой" и "межгрупповой" конкуренции. В оптимальных по температуре, освещенности и минеральному питанию условиях среды максимальная скорость роста считается зависящей только от объема клеток фитопланктона. Удельные скорости естественной смертности фитопланктона также рассчитываются в зависимости от объема клеток так, чтобы обеспечить более крупным клеткам меньший темп смертности и большую толерантность к ухудшению условий среды. Функциональная зависимость от объема клеток фитопланктона введена и в соотношения, задающие интенсивность оседания водорослей из верхнего слоя в нижний. Схема расчета потоков вещества, определяемых трофодинамическими взаимодействиями в звеньях фитопланктон - зоопланктон - детрит, основана на эмпирических зависимостях. При этом учитываются различия в рационе у разных групп зоопланктона, в коэффициентах усвояемости пищи и ее доступности. Максимальные частные (видоспецифичные) удельные рационы задаются в модели как функция средней массы представителей экологически однородной группы зоопланктона, объема пищевых частиц и температуры воды. Удельная скорость естественной смертности животных и интенсивность экскреции продуктов катаболизма, отождествляемой с тратами на обмен, задается константами, увеличивающимися с уменьшением массы зоопланктонных организмов. Интенсивность онтогенетического метаморфоза задается с учетом числа науплиальных и копеподитных стадий, объединенных в соответствующей экологически однородной группе зоопланктона, и продолжительности пребывания в ней, определяемой температурой воды.
Таким образом, данная модель интересна использованием ряда оригинальных приемов:
1. дифференциация планктонного сообщества на экологически однородные группы неразрывно связана с принципами формального описания их взаимодействий;
2. параметры, характеризующие потребление пищи, предполагаются зависящими от морфологических характеристик как хищника, так и жертвы;
3. учтена обратная связь между скоростью роста и концентрацией фитопланктона, от которой зависят лимитирующие эффекты света и биогенов;
4. при описании процессов фотосинтеза и питания животных введены температурные оптимумы.
В работе (Xu et al., 1999)рассматриваются две экологические модели, которые описывают динамику фосфора в пищевой цепи озера Чао (Китай) и исследуется влияние макрофитов на эту динамику (модель 1 - без макрофитов, модель 2 - с макрофитами). Модель 1 включает в себя следующие переменные состояния: биогенные элементы, фито-, зоопланктон, рыбы, детрит и блок, описывающий процессы в седиментах. В модель 2, помимо перечисленных переменных, включен блок, описывающий процессы в макрофитах. Показано, что макрофиты в эвтрофных озерах играют важную роль в регулировании биологической структуры озера, поскольку они, например, могут ограничивать развитие фитопланктона вследствие конкуренции за пищевые ресурсы и изменения световых условий водоема, способствовать увеличению биомассы нехищных видов рыб, exergy, structural exergy, прозрачности и отношения биомассы зоопланктона к биомассе фитопланктона.
Динамика фитопланктона включает в себя следующие процессы: рост, дыхание, осаждение, выедание зоопланктоном, естественная смертность, не связанная с трофическими взаимодействиями, вынос из водоема за счет естественного водообмена. Скорость роста фитопланктона описывается традиционно: с помощью произведения максимальной скорости роста и функций коррекции на температурные, световые условия и условия питания. В данных моделях использовалась температурная функция с оптимумом. Световое лимитирование задавалось в виде функции с насыщением, проинтегрированной по глубине. Коэффициент экстинкции определялся как линейная сумма нескольких коэффициентов экстинкции, представляющих собой поглощение света водой (без учета поглощения растворенным м взвешенным органическим веществом), фитопланктоном и макрофитами. Это выражение позволяет учитывать эффекты самозатенения на рост фитопланктона. Биогенное лимитирование роста фитопланктона задается в виде двухступенчатого процесса, причем в качестве лимитирующего биогенного элемента рассматривается только фосфор. На первой стадии этого процесса происходит увеличение содержания фосфора внутри клеток фитопланктона до определенного уровня. Только затем происходит уже рост клеток. Предполагается, что фосфорное лимитирование роста фитопланктона зависит от текущего уровня внутриклеточного содержания этого элемента, а также от минимальной клеточной квоты по фосфору. Поэтому в модели внутриклеточное содержание фосфора рассматривается как отдельная переменная состояния, которая зависит от скорости поглощения фосфора и скорости роста фитопланктона, причем скорость поглощения зависит и от внутреннего, и от внешнего содержания фосфора. Смертность фитопланктона задается в модели в виде произведения максимальной скорости смертности на корректирующую функцию, учитывающую зависимость смертности от температуры. Аналогично описываются траты на дыхание, которые связаны с метаболическими процессами. Скорость оседания фитопланктона зависит от плотности клеток и воды, размера, формы, физиологического состояния клеток, температуры и вязкости воды, скорости течения и турбулентного обмена. В данной модели скорость оседания задается как функция некоторой постоянной скорости оседания с поправкой на глубину и температуру воды. Процессы роста, отмирания макрофитов, зоопланктона и рыб, их траты на обмен описываются аналогично, но с другими коэффициентами. Для зоопланктона и рыб дополнительно учитывается возможное увеличение смертности, связанное с отравлением токсинами, выделяемыми фитопланктоном. Динамика растворимого фосфора и процессы в седиментах описываются как простые реакции первого порядка с коррекцией скоростей на температурные условия. Эта модель очень много заимствует из модели, разработанной ранее для целей управления состоянием озерных экосистем (Йоргенсен, 1985).
Дополнительная информация:
© 2001-2026 Кафедра биофизики МГУ