Биотические взаимоотношения в планктоне с участием микробиальных организмов
В работе (Porter et al., 1988) выделены следующие основные компоненты микробиальной пресноводной пищевой цепи (A - автотрофный, H - гетеротрофный, P - фаготрофный).
|
Компонент микробиальной пищевой цепи |
Трофическая форма |
|
Picoplankton (0.2 - 2 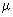 m) m) |
|
|
bacteria |
H |
|
photo and chemolithotrophs |
A |
|
coccoid cyanobacteria |
A, H |
|
small eukaryotes |
A, H |
|
Nannoplankton (2 - 20 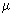 m) m) |
|
|
Flagellates |
|
|
Heterotrophic microflagellates |
H |
|
Autotrophic microflagellates |
A, H |
|
Mixotrophic flagellates |
A, H, P |
|
Ciliates |
|
|
colorless |
P, H |
|
pigmented |
P, A, H |
|
Amoebae |
P, A, H |
|
Euglenoids |
|
|
colorless |
P, H |
|
pigmented |
A, H |
|
Microzooplankton (20 - 200 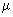 m) m) |
|
|
Ciliates |
Omnivore |
|
Rotifers |
Omnivore |
|
Nauplii |
Omnivore |
|
Macrozooplankton (200 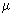 m - 2 mm) m - 2 mm) |
|
|
Cladocera |
Omnivore |
|
Copepods |
Omnivore |
Степень развития микробиальных организмов определяется, с одной стороны, трофическими ресурсами, которые поставляет фитопланктон, с другой - выеданием метазойным планктоном, то есть регулируется в рамках трофической сети “снизу” и “сверху”. Бактериопланктон активно выедается многими простейшими и тонкими фильтраторами зоопланктона (влияние “сверху”). В то же время его обилие часто лимитируется растворенным органическим веществом и, таким образом, тесно зависит от углеродного метаболизма фитопланктона (влияние “снизу”), поэтому увеличение или снижение биомассы и продукции фитопланктона приводит к аналогичному отклику бактерий (Currie, Kalff, 1984; Currie, Kalff, 1984; Beaver, Crisman, 1989; Billen et al., 1990). Подтверждением влияния “снизу” служит положительная корреляция численности бактерий и их активности с биомассой фитопланктона и его продукцией (Бульон, Павельева, 1998; Bird, Kalff, 1984; Sondergaard et al., 1985; Currie, 1990).
К биотическим факторам, регулирующим обилие микробиальных организмов “сверху”, относится выедание кормовых частиц планктонными веслоногими и ветвистоусыми ракообразными с фильтрационным типом питания (Монаков, 1976; Гутельмахер и др., 1988; Монаков, 1998; Бульон и др, 1999). У веслоногих рачков (Copepoda) ток воды с пищевой взвесью, создаваемый вибрацией ротовых конечностей, направлен в околоротовое пространство, ограниченное щетинками максилл и максиллярных ног. Расстояние между щетинками у отдельных видов и возрастных групп пресноводных копепод изменяется в границах от 1,7 до 15 мкм. У ветвистоусых рачков (Cladocera) фильтрационный аппарат образуют щетинки торакальных конечностей. Расстояние между ними варьирует в основном от 0,2 до 4 мкм. Именно частота расположения щетинок определяет минимальные размеры потребляемой пищи. По густоте щетинок в фильтрационных аппаратах все рачки принято делить на “грубых” и “тонких” фильтраторов. Концепция микробиальной “петли” впервые разработана на примере морских экосистем, где в зоопланктоне преобладают копеподы. Каланоиды как грубые фильтраторы и циклопоиды как факультативные хищники фактически не потребляют такие мелкие организмы, как бактерии и микрофлагелляты. Поэтому при доминировании копепод заметно усиливается функция микробиального сообщества. Кладоцеры, например Ceriodaphnia и Daphnia, - основные бактериофаги среди планктонных ракообразных. Для них характерен широкий спектр питания. Потребляя как автотрофный, так и гетеротрофный микропланктон размером от 1 до 50 мкм, кладоцеры разрушают микробиальную “петлю” (Pace et al., 1990).
При сходных биомассах зоопланктона обилие фитопланктона, как правило, связано обратной зависимостью со средними размерами планктонных животных. Это объясняется тем, что пресс на фитопланктон оказывает в первую очередь крупный зоопланктон. Выедая водоросли, зоопланктон косвенно снижает численность микробиальных организмов - подрывает их энергетическую базу.
Концепция микробиальной трофической сети обычно рассматривает только планктонных ракообразных как важнейший фактор, регулирующий обилие простейших и бактерий “сверху”. В результате из поля зрения этой концепции выпали другие активные консументы, например, коловратки из рода Asplanchna (Бульон и др., 1999). Между тем аспланхна является одним из массовых представителей крупного зоопланктона и, следовательно, играет заметную роль в регулировании структуры планктонного сообщества. Считается, что широко распространенные в пресных водах виды рода Asplanchna относятся к полифагам, однако нормально существовать могут только на животной пище. Соотношение растительной и животной пищи в их рационе зависит от вида и трофических условий в водоеме. Как менее специализированный хищник рассматривается A. priodonta. Из животной пищи в ее рацион входят мелкие коловратки и простейшие. Из растительной пищи, которая нередко преобладает в их рационе, - одиночные крупные клетки и колонии водорослей.
Функция микробиальной “петли” в планктонном сообществе обсуждается в литературе, как правило, лишь на качественном уровне. В работе (Бульон и др., 1999) представлены одни из первых количественные данные о непосредственном влиянии A. priodonta на структуру микробиального сообщества и на обилие тех его компонентов, которые могут быть жертвой этого факультативного хищника. Было показано, что при сильном стрессе отношение биомассы зоопланктона к биомассе фитопланктона приближалось к единице, при слабом - это отношение снижалось до 0,06, при умеренном - составляло около 0,2. В результате удалось выявить обратную зависимость обилия микробиальных организмов (бактерий и простейших) от отношения 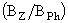 , которую предлагается использовать в качестве индекса биотических взаимодействий в планктоне. Реакция на соотношение биомасс консументов и автотрофных организмов была наиболее выраженной у простейших. Бактерии в этом отношении были более консервативны. Если воздействие на бактерий “сверху” и оказывалось, то, очевидно, не со стороны A. priodonta, а со стороны кладоцер и в меньшей степени со стороны цилиат. Однако выедание бактерий, по-видимому, быстро компенсировалось позитивным влиянием на них трофических ресурсов, поставляемых фитопланктоном и фондом РОВ, формирующимся в периоды “избыточного” продуцирования органического вещества водорослями. Бактериопланктон всегда занимал лидирующее положение в потоке органического вещества и рассеивании энергии, что в значительной мере определяло конфигурацию, степень выраженности и функциональную значимость микробиальной “петли”. При сильном прессе метазойного планктона активно функционирует только одна ее ветвь: фитопланктон - бактерии. Другая ее ветвь (фитопланктон - простейшие) находится в сильно редуцированном состоянии. Можно сказать, что в данном случае микробиальная “петля” по своей конфигурации асимметрична. При слабом прессе метазойного планктона активно функционируют обе ветви микробиальной “петли”, то есть она принимает симметричную форму. В зависимости от степени выраженности микробиальной “петли” соотношение продукции метазойного планктона и продукции фитопланктона варьировало от 4-5 до 20% и только при некоторых средних условиях, при умеренном развитии микробиальных микроорганизмов, это соотношение близко к 10%, то есть к “классическому” значению эффективности переноса энергии между смежными трофическими уровнями (Иванова, 1985).
, которую предлагается использовать в качестве индекса биотических взаимодействий в планктоне. Реакция на соотношение биомасс консументов и автотрофных организмов была наиболее выраженной у простейших. Бактерии в этом отношении были более консервативны. Если воздействие на бактерий “сверху” и оказывалось, то, очевидно, не со стороны A. priodonta, а со стороны кладоцер и в меньшей степени со стороны цилиат. Однако выедание бактерий, по-видимому, быстро компенсировалось позитивным влиянием на них трофических ресурсов, поставляемых фитопланктоном и фондом РОВ, формирующимся в периоды “избыточного” продуцирования органического вещества водорослями. Бактериопланктон всегда занимал лидирующее положение в потоке органического вещества и рассеивании энергии, что в значительной мере определяло конфигурацию, степень выраженности и функциональную значимость микробиальной “петли”. При сильном прессе метазойного планктона активно функционирует только одна ее ветвь: фитопланктон - бактерии. Другая ее ветвь (фитопланктон - простейшие) находится в сильно редуцированном состоянии. Можно сказать, что в данном случае микробиальная “петля” по своей конфигурации асимметрична. При слабом прессе метазойного планктона активно функционируют обе ветви микробиальной “петли”, то есть она принимает симметричную форму. В зависимости от степени выраженности микробиальной “петли” соотношение продукции метазойного планктона и продукции фитопланктона варьировало от 4-5 до 20% и только при некоторых средних условиях, при умеренном развитии микробиальных микроорганизмов, это соотношение близко к 10%, то есть к “классическому” значению эффективности переноса энергии между смежными трофическими уровнями (Иванова, 1985).
Структуру биотических взаимодействий в планктоне при участии микробиальных организмов в первом приближении можно описать следующим образом:
·биомасса фитопланктона строго контролируется метазойным планктоном;
·выедание метазойным планктоном и трофическая стимуляция фитопланктоном - ключевые факторы, регулирующие обилие бактерий и простейших “сверху” и “снизу”;
·метазойный планктон принимает также опосредованное участие в формировании гетеротрофного микробиального сообщества - через редукцию трофических ресурсов, поставляемых фитопланктоном.
Трансформация вещества и энергии через микробиальное сообщество. В пределах микробиальной “петли” потоки углерода и биогенных элементов тесно связаны между собой и взаимно согласованы (сопряжены). Динамическое поведение внутри этой структуры определяется взаимодействием трех основных процессов: образованием РОВ фитопланктоном и утилизацией его бактериями (комменсализм), конкуренцией за пищу между бактериями и фитопланктоном, хищничеством между флагеллятами, цилиатами и микрозоопланктоном, в результате чего формируется цепь обратной связи, пополняющая запасы биогенных веществ и РОВ (Stockner, Porter, 1988).
Легкоусвояемое органическое вещество (РОВ) водоемов формируется за счет прижизненных и посмертных выделений организмов. Основной его потребитель - фито- и бактериопланктон. Несмотря на то, что энергию водоросли получают за счет фотосинтеза, хорошо известна и их способность потреблять ОВ. Последние в этом случае становятся конкурентами бактерий (Гусев, Никитина, 1979; Кузьменко, 1981; Сиренко, Козицкая, 1988). Однако участие фитопланктона в разрушении РОВ чаще всего не рассматривается и считается, что основную роль в этом процессе играют бактерии. Известно, что РОВ низкой молекулярной массы неодинаково потребляются клетками организмов. Низкомолекулярные соединения потребляются водорослями и бактериями непосредственно, тогда как высокомолекулярные предварительно должны быть гидролизованы экзоферментами. Следовательно, спектр организмов-потребителей этого РОВ должен быть значительно уже. Водоросли неспособны экскретировать в среду литические ферменты (в частности, протеазы). Основную роль в ферментативном гидролизе ОВ играют бактерии, которые оказывают “помощь” водорослям в их гетеротрофии. Таким образом, несмотря на интенсивное потребление РОВ водорослями, гетеротрофная активность последних в значительной степени зависит от жизнедеятельности бактерий (Садчиков, 2002). Утилизация высокомолекулярных соединений водорослями осуществляется только в присутствии бактерий. Именно бактерии выделяют в водную среду литические экзоферменты, которые разрушают высокомолекулярные фракции РОВ и подключают к процессу дальнейшего разрушения ОВ и других гидробионтов, которые способны трансформировать только низкомолекулярные соединения.
Микробиальная “петля” и линейная пищевая цепь представляют собой сложную сеть трофических взаимодействий (своеобразный природный триггерный механизм), который управляется влиянием “снизу” и “сверху”, а также абиотическими факторами, в особенности температурными условиями. Продукция фитопланктона частично ассимилируется и бактериями, и простейшими, частично трансформируется через цепь бактерии - жгутиковые - цилиаты к метазойному планктону. Хищничество гетеротрофных флагеллят на бактериях служит важной ступенью в трансформации органического углерода к крупному зоопланктону. Цилиаты также относятся к активным бактериофагам и к консументам по отношению к мелким флагеллятам. Они обеспечивают еще одну связь с верхним трофическим уровнем. Эффективность переноса энергии между компонентами микробиальной “петли” составляет от 10 до 40% (Sheldon et al., 1986; Madoni et al., 1990; Weisse et al., 1990). Следовательно, чем больше компонентов участвует в потоке вещества, тем выше общий метаболизм сообщества и тем интенсивнее рециклинг биогенных элементов. Эффективность трансформации вещества через микробиальную сеть зависит от степени ее сложности и характера ее взаимодействия с классической пищевой цепью. В свою очередь характер этого взаимодействия в значительной степени контролируется консументами, представляющими классическую пищевую цепь (Porter et al., 1988).
Тонкие фильтраторы, к которым относятся кладоцеры, могут использовать неагрегированный бактериопланктон как один из источников органического углерода. Виды рода Daphnia считаются ключевыми компонентами планктонных сообществ. Они потребляют организмы, размерный диапазон которых охватывает несколько трофических уровней: гетеротрофный и автотрофный пикопланктон, инфузорий, нанофитопланктон и микрозоопланктон (Stockner, Porter, 1988; Pace et al., 1990; Pace, Vaque, 1994; Vezina, Pace, 1994).
В отличие от них копеподы - грубые фильтраторы ограничиваются выеданием в основном только крупных жертв (Porter et al., 1988; Nagata et al., 1996), поэтому между копеподами и бактериями должны быть дополнительные звенья - гетеротрофные флагелляты, цилиаты и микрозоопланктон (мелкие коловратки), которые активно потребляют бактерий (Sorokin, 1981; Pace, 1986). Однако с появлением дополнительных трофических звеньев снижается эффективность переноса органического углерода от продуцентов в “классическую” пищевую цепь (Stockner, Porter, 1988).
Таким образом, кладоцеры, редуцируя микробиальную “петлю”, сокращают трофическую цепь и тем самым повышают эффективность трансформации органического углерода от автотрофных организмов к верхнему трофическому уровню. В присутствии копепод трофическая цепь удлиняется, в результате чего эффективность перехода органического углерода к верхнему трофическому уровню снижается. Так как деструкция органического вещества на всех трофических уровнях сопряжена с регенерацией биогенных элементов (в частности, фосфора), рециклинг последних ускоряется в системах баз кладоцер (Pace et al., 1990).
Планктоноядные рыбы, выедая крупных кладоцер, также снижают эффективность переноса энергии от первичных продуцентов, так как значительная ее доля рассеивается при дыхании микробиальных организмов, освобожденных от чрезмерного пресса консументов. Важно подчеркнуть, что снижение (или повышение) эффективности трансформации органического углерода в планктонном сообществе сопровождается ускорением (или замедлением) рециклинга фосфора. Происходит как бы разобщение потока органического углерода и круговорота фосфора при доминировании в планктоне как кладоцер, так и копепод. Можно предположить, что только при сбалансированной активности этих двух ключевых компонентов зоопланктона структура трофической сети приобретает переходную форму между классической линейной пищевой цепью и цепью, усложненной микробиальной “петлей”. Итак, поток органического вещества от первичных продуцентов к метазойному планктону может идти одновременно как через микробиальную трофическую сеть, так и по короткому пути, минуя микробиальную трофическую сеть, разветвляясь по обоим каналам в том или ином соотношении.
В работе (Weisse et al., 1990) отмечено, что поскольку микробиальная пищевая цепь представляет собой достаточно динамичную структуру то, следовательно, для оценки энергетических потоков важное значение имеет выбор соответствующего временного масштаба. Например, нельзя рассматривать и анализировать структуру микробиальной “петли” в терминах годовых биомасс или накопленной продукции.
Связь между активностью микробиального сообщества и трофическим статусом озер. Согласно распространенной точке зрения, вклад микробиальных процессов возрастает вдоль трофического градиента озер: от эвтрофных к олиготрофным системам (Porter et al., 1988), причем в олиготрофных озерах микробиальные процессы достигают наибольшего значения в период летней стратификации, когда в эпилимнионе истощаются биогенные элементы. Выше было указано, что только микробиальные организмы способны утилизировать следовые количества биогенных элементов и ускорять их рециклинг, поддерживая тем самым автотрофный синтез органического вещества.
Таким образом, анализ литературы по структурно-функциональным связям в трофических цепях позволяет сформулировать следующие основные выводы (Бульон, 2002).
1. Концепция микробиальной “петли” в отличие от классической концепции линейной пищевой цепи делает акцент на активном участии микробиальных организмов (бактерий и простейших) в потоке вещества и энергии в планктонном сообществе. Микробиальная трофическая сеть - неотъемлемая ступень в трансформации первичной продукции, в ней аккумулируются и подвергаются регенерации биогенные элементы.
2. Благодаря ее функционированию ускоряется рециклинг биогенных элементов и предотвращается их уход из фотической зоны, что способствует сохранению первичной продукции на относительно высоком уровне даже при дефиците минеральных форм фосфора. Однако при сильно выраженной микробиальной “петле” из-за удлинения и усложнения трофической сети увеличивается доля рассеиваемой энергии, в результате чего продукция метазойного планктона снижается относительно первичной продукции. При слабо выраженной микробиальной “петле” рециклинг биогенных элементов замедляется, но возрастает эффективность переноса энергии первичной продукции к метазойному планктону. Налицо “разобщение” потока энергии, заключенной в органическом углероде, и круговорота биогенных элементов (в первую очередь фосфора) как при ярко выраженной микробиальной “петле”, так и при слабом ее проявлении.
3. Поскольку степень выраженности микробиальной “петли” определяется биотическими воздействиями”снизу” и “сверху”, то можно прийти к следующему парадоксальному выводу. Если регулирование “сверху” преобладает над встречным воздействием, планктонное сообщество функционирует по укороченной схеме фитопланктон - зоопланктон. Благодаря снижению трат на обмен сообщество становится более экономичным в отношении энергии, но при этом пребывает в неустойчивом состоянии, так как зоопланктон, интенсивно выедая не только продукцию, но и часть продуцирующей биомассы фитопланктона, подрывает собственную кормовую базу. Если же превалирует регулирование “снизу”, планктонное сообщество как трофическая сеть усложняется за счет интенсивного развития микробиальных организмов, приобретает тем самым повышенную устойчивость, но расплачивается за это дополнительными тратами энергии на дыхание.
4. Не решен окончательно вопрос о том, являются ли микробиальные процессы следствием или, напротив, детерминантом структуры планктонного сообщества. Вероятнее всего микробиальная трофическая сеть и планктонное сообщество в целом оказывают обоюдное влияние. С одной стороны, обратные трофические воздействия по типу хищник - жертва приводят к редукции численности микробиальных организмов, с другой - выделяемые консументами биогенные элементы стимулируют развитие своих жертв. Микробиальные организмы в свою очередь “шунтируют” пищевую цепь, если фитопланктон представлен формами, слабо потребляемыми зоопланктоном.
5. Разработка концепции микробиальной трофической сети еще далека от завершения. Одна из ее слабых сторон состоит в том, что продукция фитопланктона рассматривается как основной источник энергии для гетеротрофных микроорганизмов. В действительности для них существуют два источника энергии: 1) остаток продукции фитопланктона после потребления ее зоопланктоном в сумме с неусвоенной зоопланктоном пищей; 2) аллохтонное органическое вещество в сумме с органическим веществом, ранее синтезированным фитопланктоном и аккумулированным в виде детрита. Эти два источника формируют так называемую первичную энергию, которую необходимо учитывать при моделировании краткосрочных энергетических балансов в локальных планктонных сообществах.
6. В концепции микробиальной трофической сети должна быть учтена роль детрита и растворенного органического вещества, так как заключенная в них энергия с той или иной эффективностью ассимилируется гетеротрофными организмами, что делает их менее зависимыми от флуктуации первичной продукции.
Дополнительная информация:
© 2001-2026 Кафедра биофизики МГУ