Для изучения условий формирования неоднородностей фитопланктона важно определить пространственно-временные масштабы взаимодействия физических и биологических процессов в водных экосистемах (Картушинский, 1998). Воздействие гидродинамических факторов на фотосинтез фитопланктона не однозначно. Они могут как усиливать, так и ослаблять продукционные процессы. На основе обобщения экспериментальных и теоретических исследований в фитопланктонном сообществе выделено четыре основных явления, возникающих под действием перемешивания (Крейман и др., 1992): 1) усиление обменных процессов между зоной питания клетки и окружающей водной средой; 2) ослабление конкуренции между организмами за световые и питательные ресурсы; 3) изменение скорости оседания; 4) механическое перераспределение водорослей и биогенных элементов в пределах водоема и отдельных слоев. Эти явления отражают положительное влияние турбулентного перемешивания на продуктивность фитопланктона. Вместе с тем, при интенсивном турбулентном перемешивании в штормовых условиях отмечается практически полное подавление процесса фотосинтеза. Недооценка влияния турбулентности на фотосинтез водорослей может приводить к ошибкам (до 200%) в расчетах образования ОВ, поскольку турбулентность оказывает влияние практически на все важнейшие процессы, определяющие скорости первичного продуцирования ОВ в природных водоемах.
Характерная особенность вертикальной структуры морей, озер и водохранилищ - наличие в них верхнего перемешанного слоя. Как показывают фактические данные, в одних случаях вертикальное распределение фитопланктона в пределах этого слоя может быть практически однородным, в других - профили концентрации водорослей обнаруживают один или несколько экстремумов. Максимумы и минимумы на вертикальных профилях концентрации фитопланктона обычно связаны с освещенностью, концентрацией биогенных элементов, качественным и количественным развитием фито- и зоопланктонного сообществ. Однородное распределение - результат выравнивания концентрации водорослей в верхнем слое за счет механического перераспределения его турбулентными вихрями. Показателем неоднородности вертикального профиля концентрации фитопланктона может служить, например, коэффициент вариации: 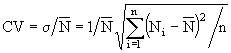 , где
, где ![]() - среднеквадратичное отклонение концентрации фитопланктона,
- среднеквадратичное отклонение концентрации фитопланктона, ![]() - средняя концентрация водорослей в перемешанном слое толщиной Н,
- средняя концентрация водорослей в перемешанном слое толщиной Н, ![]() - измеренная концентрация водорослей на глубине z, n - количество эмпирических точек, по которым строится вертикальный профиль.
- измеренная концентрация водорослей на глубине z, n - количество эмпирических точек, по которым строится вертикальный профиль.
Для количественной характеристики интенсивности турбулентности, с которой связаны особенности вертикального распределения концентрации фитопланктона в верхнем слое озера, может быть использовано потоковое число Ричардсона, являющееся интегральным показателем интенсивности турбулентного перемешивания в слое ![]() :
: ![]() , где
, где ![]() - перепад плотности в термоклине,
- перепад плотности в термоклине, ![]() - скорость трения на водной поверхности, которая рассчитывается по данным метеорологических измерений в приводном слое атмосферы. На основе статистической обработки для условий стратифицированного водоема в работе (Крейман и др., 1992) была получена формула
- скорость трения на водной поверхности, которая рассчитывается по данным метеорологических измерений в приводном слое атмосферы. На основе статистической обработки для условий стратифицированного водоема в работе (Крейман и др., 1992) была получена формула ![]() , связывающая вертикальное распределение фитопланктона с интенсивностью турбулентного перемешивания в верхнем слое. Можно выделить три диапазона чисел
, связывающая вертикальное распределение фитопланктона с интенсивностью турбулентного перемешивания в верхнем слое. Можно выделить три диапазона чисел ![]() , которые соответствуют различным режимам поведения водорослей в турбулизированной водной среде. При
, которые соответствуют различным режимам поведения водорослей в турбулизированной водной среде. При ![]() коэффициент вариации характеризуется малыми значениями и существенной зависимостью от
коэффициент вариации характеризуется малыми значениями и существенной зависимостью от ![]() , что отражает преобладающее влияние турбулентности на перераспределение фитопланктона. Так как при интенсивном турбулентном перемешивании фитопланктон практически лишен возможности длительное время находиться в условиях оптимального обитания, то в данном диапазоне чисел
, что отражает преобладающее влияние турбулентности на перераспределение фитопланктона. Так как при интенсивном турбулентном перемешивании фитопланктон практически лишен возможности длительное время находиться в условиях оптимального обитания, то в данном диапазоне чисел ![]() можно ожидать подавления фотоснтеза водорослей турбулентностью. При
можно ожидать подавления фотоснтеза водорослей турбулентностью. При ![]() снижается турбулизация охваченного перемешиванием водного слоя, CV в меньшей степени зависит от числа Ричардсона, что свидетельствует об ослаблении действия “турбулентного пресса” на перераспределение фитопланктона. При относительно слабой интенсивности турбулентности в верхнем водном слое (
снижается турбулизация охваченного перемешиванием водного слоя, CV в меньшей степени зависит от числа Ричардсона, что свидетельствует об ослаблении действия “турбулентного пресса” на перераспределение фитопланктона. При относительно слабой интенсивности турбулентности в верхнем водном слое (![]() ) величины CV перестают зависеть от
) величины CV перестают зависеть от ![]() . Поскольку CV косвенно характеризует способность водорослей к самостоятельному перераспределению в пределах трофогенной зоны, то развитие фитопланктона полностью определяется условиями освещенности и насыщенностью водной среды биогенными веществами, а также межвидовыми отношениями в фитопланктонном сообществе. Вероятно, именно при
. Поскольку CV косвенно характеризует способность водорослей к самостоятельному перераспределению в пределах трофогенной зоны, то развитие фитопланктона полностью определяется условиями освещенности и насыщенностью водной среды биогенными веществами, а также межвидовыми отношениями в фитопланктонном сообществе. Вероятно, именно при ![]() слабая турбулентность может оказывать положительное воздействие на фотосинтез водорослей.
слабая турбулентность может оказывать положительное воздействие на фотосинтез водорослей.
Для полностью перемешанных водоемов и, в силу этого, однородных по плотности степень развития турбулентности характеризуется числом Рейнольдса. На основе лабораторных экспериментов была установлена зависимость безразмерной концентрации клеток ![]() (N - концентрация клеток фитопланктона в эксперименте с перемешиванием,
(N - концентрация клеток фитопланктона в эксперименте с перемешиванием, ![]() - в контроле) от числа Re:
- в контроле) от числа Re: ![]() . И здесь можно выделить три диапазона чиел Re. Первый режим характеризуется малыми числами Re (<60) и значениями
. И здесь можно выделить три диапазона чиел Re. Первый режим характеризуется малыми числами Re (<60) и значениями ![]() , что говорит о положительном влиянии турбулентности малой интенсивности на фотосинтез фитопланктона в экспериментальном объеме. Во втором режиме достаточно резко убывают величины
, что говорит о положительном влиянии турбулентности малой интенсивности на фотосинтез фитопланктона в экспериментальном объеме. Во втором режиме достаточно резко убывают величины ![]() : от 1,2-1,3 при Re = 60, до 0,2 при Re = 120. Это указывает на быстрое ингибирование развития водорослей усиливающимся перемешиванием. Третий режим (
: от 1,2-1,3 при Re = 60, до 0,2 при Re = 120. Это указывает на быстрое ингибирование развития водорослей усиливающимся перемешиванием. Третий режим (![]() ) свидетельствует о фотосинтезе, заметно подавленном перемешиванием (концентрация клеток при больших Re в среднем составляет лишь около 20% концентрации в контроле) и маломеняющемся при росте числа Re до максимальной для этих опытов ее величины (3500).
) свидетельствует о фотосинтезе, заметно подавленном перемешиванием (концентрация клеток при больших Re в среднем составляет лишь около 20% концентрации в контроле) и маломеняющемся при росте числа Re до максимальной для этих опытов ее величины (3500).
Наиболее устойчивый характер взаимодействий гидрофизических процессов с фитопланктоном наблюдается в металимнионе или высокоградиентных слоях (Картушинский, 1998). При увеличении ветровой нагрузки существенно меняется структура распределения концентрации хлорофилла “а” за счет изменений гидрологической структуры водного объекта. При слабых ветровых нагрузках и небольшой адвективной составляющей течения, клетки фитопланктона накапливаются в верхнем фотосинтетическом слое. Накопление зависит от активного формирования поля температуры и плотности. Чем слабее выражена плотностная стратификация, тем больше вертикальный масштаб неоднородного распределения фитопланктона. Установлено, что температурный фактор и плотностная стратификация вод формируют ограничительный уровень воздействия на фитопланктон в условиях турбулентного перемешивания и потока тепла совместно с динамическим воздействием ветра на водную поверхность. Периодичность скорости ветра и интенсивность ветрового касательного напряжения в течение суток приводит к эффекту расслоения водных масс и формированию плотностных градиентов, препятствующих однородному распределению клеток фитопланктона по всей толще вод. Световой фактор обостряет различия между слоями воды по величине биомассы из-за эффекта фотоингибирования в верхних слоях и затенения на компенсационных глубинах, в зависимости от времени суток. Динамический фактор, связанный с вертикальными движениями вод, регулирует поступление биогенных элементов из глубоких слоев и изменяет структуру распределения биомассы фитопланктона в течение суток.
Таким образом, турбулентность представляет собой сильный лимитирующий фактор. Турбулентное перемешивание в степени не меньшей, чем другие абиотические характеристики среды, необходимо учитывать при расчетах пространственного распределения и скорости роста фитопланктона.
Важность турбулентного перемешивания для поддержания клеток диатомовых водорослей в верхнем фотическом слое показана в работе (Гранин и др., 1999). Клетки Aulacoseira baicalensis не обладают физиологическим механизмом, который обеспечивал бы их положительную плавучесть. Из-за тяжелого кремнистого панциря их плотность достигает 1,1 г/см3, и они погружаются под действием силы тяжести со средней скоростью 3-4 метра в сутки. Известно, что развитие A. baicalensis происходит зимой, обычно с конца февраля до апреля, подо льдом. Поэтому решающее значение для поддержания концентрации, достаточной для вегетативного размножения A. baicalensis в прилегающем ко льду слое воды, имеют два фактора: скорость деления клеток (зависящая от освещенности) и интенсивность турбулентности, возвращающая тонущие клетки из нижних слоев воды в фотический слой. В данной работе описываются физические характеристики прилегающего ко льду тонкого слоя воды, в котором происходит половое, а затем и вегетативное размножение A. Baicalensis. Известно, что конвекция и турбулентность возникают на Байкале в подледный период за счет потока плавучести, вызванного изменением плотности при объемном поглощении солнечной энергии. Этот механизм начинает интенсивно работать в начале или середине марта и постепенно охватывает слой толщиной 20 - 50 метров. Однако он не является определяющим для сохранения популяции A. Baicalensis в подледный период, так как половое размножение происходит раньше, в то время, когда поток солнечной энергии минимален и происходит нарастание толщины льда.
В работе описывается другой механизм формирования турбулентности в тонком слое воды, непосредственно примыкающим ко льду. Этот механизм работает именно тогда, когда толщина льда нарастает. Поток плавучести обеспечивается отдачей солей при превращении воды в лед. Повышение минерализации при отдаче солей намерзающим льдом вызывает повышение плотности воды в прилегающем ко льду слое и приводит к возникновению конвекции. Можно полагать, что конвекция за счет осолонения имеет место при ![]() , где
, где ![]() и
и ![]() - разности минерализации и температуры по вертикали,
- разности минерализации и температуры по вертикали, ![]() - удельный коэффициент температурного расширения,
- удельный коэффициент температурного расширения, ![]() - удельное изменение плотности, вызванное изменением минерализации
- удельное изменение плотности, вызванное изменением минерализации ![]() . Оценки показывают, что при
. Оценки показывают, что при ![]() = 10 мг/л и средней минерализации воды Байкала
= 10 мг/л и средней минерализации воды Байкала ![]() 96 мг/л
96 мг/л ![]() 0,15°С. Это означает, что в середине марта конвекция, вызванная осолонением прилегающего ко льду слоя, может проникнуть на глубину залегания изотермы 0,15°С, то есть около 2 метров. К концу марта, когда под воздействием возрастающей инсоляции температура прилегающего ко льду слоя воды возрастает, а скорость намерзания льда падает, глубина проникновения конвекции, вызванной осолонением, уменьшается, а затем, когда лед начинает снизу подтаивать, процесс прекращается. В период прогрева, который начинается в середине марта, конвекция, обусловленная объемным поглощением солнечной радиации, проникает до глубины 50 м.
0,15°С. Это означает, что в середине марта конвекция, вызванная осолонением прилегающего ко льду слоя, может проникнуть на глубину залегания изотермы 0,15°С, то есть около 2 метров. К концу марта, когда под воздействием возрастающей инсоляции температура прилегающего ко льду слоя воды возрастает, а скорость намерзания льда падает, глубина проникновения конвекции, вызванной осолонением, уменьшается, а затем, когда лед начинает снизу подтаивать, процесс прекращается. В период прогрева, который начинается в середине марта, конвекция, обусловленная объемным поглощением солнечной радиации, проникает до глубины 50 м.
Поток плавучести J, вызванный осолонением, может быть оценен подобно тому, как оценивается поток плавучести при испарении. Если m есть скорость намерзания льда, то ![]() . Поскольку в подледный период поток кинетической энергии, обусловленный трением ветра, отсутствует, можно предположить, что скорость диссипации турбулентной энергии приблизительно равна потоку плавучести, вызванному осолонением. При минерализации воды 96 мг/л и льда 20 мг/л и скорости намерзания льда около 1см/сутки поток плавучести, а следовательно, и скорость диссипации турбулентной энергии составит
. Поскольку в подледный период поток кинетической энергии, обусловленный трением ветра, отсутствует, можно предположить, что скорость диссипации турбулентной энергии приблизительно равна потоку плавучести, вызванному осолонением. При минерализации воды 96 мг/л и льда 20 мг/л и скорости намерзания льда около 1см/сутки поток плавучести, а следовательно, и скорость диссипации турбулентной энергии составит ![]() 6×10-11м2/с3. Вертикальный масштаб конвекции h » 1м. Отсюда коэффициент вертикального обмена
6×10-11м2/с3. Вертикальный масштаб конвекции h » 1м. Отсюда коэффициент вертикального обмена ![]() м2 c-1.
м2 c-1.
Известно уравнение, описывающее временную изменчивость вертикального распределения фитопланктона при его осаждении с учетом размножения в турбулентной среде: ![]() , где
, где ![]() - скорость роста концентрации клеток фитопланктона, V - скорость опускания фитопланктона в отсутствие турбулентности. Решение уравнения зависит от знака выражения
- скорость роста концентрации клеток фитопланктона, V - скорость опускания фитопланктона в отсутствие турбулентности. Решение уравнения зависит от знака выражения ![]() . Если эта величина положительна, то концентрация фитопланктона растет. Оценки показывают, что при найденном коэффициенте вертикального обмена и благоприятных условиях освещенности концентрация фитопланктона в верхнем слое не будет убывать при
. Если эта величина положительна, то концентрация фитопланктона растет. Оценки показывают, что при найденном коэффициенте вертикального обмена и благоприятных условиях освещенности концентрация фитопланктона в верхнем слое не будет убывать при ![]() » 15-20 суток.
» 15-20 суток.
Теоретические и эмпирические исследования и выводы планктонной трофодинамики обычно основаны на рассмотрении изменений относительной плотности организмов системы “хищник - жертва”. В работе (Rothschild, Osborn, 1988) показано, что для оценок пищевых потребностей организмов, изучения механизмов пищевого поведения, энергетики, образования и рассеивания пятен планктона, помимо плотности, необходимо учитывать также воздействие мелкомасштабной турбулентности. Например, хищник может использовать турбулентность для того, чтобы поймать отдельную жертву, и при этом не расходовать на перемещение свою собственную энергию. Можно также предположить, что хищники и жертвы регулируют свое положение в водной толще и используют турбулентное движение как для захвата пищи, так и для убегания от хищника. Биологически важные эффекты воздействия турбулентности могут быть обнаружены только на очень малых пространственных масштабах. В этом диапазоне турбулентность является однородной и изотропной, а скорости движения воды сравнимы со скоростью движения планктонных организмов. Значения средней скорости турбулентного движения уменьшаются по мере уменьшения пространственного масштаба. На организменном уровне турбулентность оказывает влияние на трофические взаимодействия, что было показано как экспериментально, так и путем численного моделирования (Andrews, 1983). Это влияние может быть как непосредственным, например, приводить к увеличению частоты встреч между хищниками и их жертвами (Rothschild, Osborn, 1988; Evans, 1989; Yamazaki et al., 1991), так и непрямым, например, в результате изменения качества пищи (Kiorboe, 1993). Более ранние эксперименты в микрокосмах показали, что мелкомасштабная турбулентность (< 1 см), кроме влияния на пищевые цепи, также способна ускорять развитие морских веслоногих ракообразных (Oviatt, 1981; Alcaraz et al., 1988; Saiz, Alcaraz, 1991), увеличивать скорости экскреции веществ (Saiz, Alcaraz, 1992), влиять на активность копепод и их поведение (Saiz, Alcaraz, 1992). Экспериментальные исследования показали, что мелкомасштабная турбулентность влияет на скорость метаболических процессов у копепод вследствие повышения их двигательной активности. Однако надо иметь в виду, что в поведенческих экспериментах исследователь имеет дело с короткими (порядка нескольких минут) сериями событий, в то время как расчеты скоростей питания и метаболических выделений требуют гораздо более длительных экспериментов: от 12 до 24 часов. Вследствие этого, возможная связь между поведением копепод, скоростями метаболических процессов и мелкомасштабной турбулентностью требует более тщательных и длительных исследований.
В работе (Alcaraz et al., 1994) приводятся результаты исследований, которые позволяют говорить о том, что мелкомасштабная турбулентность напрямую влияет на метаболизм рачкового зоопланктона (как пресноводного, так и морского) и что это воздействие происходит на тех же временных масштабах, на которых имеет место воздействие турбулентности на поведение (плавание, питание, реакции избегания). Было показано, что частота сердцебиений (HBR) является хорошим устойчивым признаком, позволяющим исследовать дыхание организмов и их метаболизм. Ускорение процессов при воздействии турбулентности связано со стимуляцией антеннальных рецепторов. Скорость деформации жидкости воспринимается сенсорными щетинками и преобразуется в нервные импульсы (Tiselius, Jonsson, 1990). Это объяснение согласуется с результатами электрофизиологических и морфологических исследований антеннальных щетинок копепод (Yen et al., 1992). Антеннальные рецепторы являются чувствительными органами к механическим перемещением и колебаниям на субмикронном уровне. Даже очень незначительные скорости потока ![]() могут вызывать ответную реакцию организма. У разных видов рачкового зоопланктона число щетинок, их длина и эластичность довольно сильно различаются. Все это приводит к наблюдаемым различиям в увеличении HBR при воздействии турбулентности. Полученные результаты позволяют говорить о том, что мелкомасштабная турбулентность оказывает стимулирующее воздействие на активность рачкового зоопланктона, как морского, так и пресноводного. Это воздействие аналогично воздействию температуры. Было показано, что увеличение турбулентности и повышение температуры приводит к аллометрическому повышению темпов развития, скоростей питания и экскреции, HBR и скорости дыхания у копепод.
могут вызывать ответную реакцию организма. У разных видов рачкового зоопланктона число щетинок, их длина и эластичность довольно сильно различаются. Все это приводит к наблюдаемым различиям в увеличении HBR при воздействии турбулентности. Полученные результаты позволяют говорить о том, что мелкомасштабная турбулентность оказывает стимулирующее воздействие на активность рачкового зоопланктона, как морского, так и пресноводного. Это воздействие аналогично воздействию температуры. Было показано, что увеличение турбулентности и повышение температуры приводит к аллометрическому повышению темпов развития, скоростей питания и экскреции, HBR и скорости дыхания у копепод.
Дополнительная информация:
© 2001-2026 Кафедра биофизики МГУ